Overview
- The Baldwin effect is the hypothesis that learned behaviours or phenotypic plasticity can guide evolutionary change by allowing organisms to survive in new environments through behavioural accommodation, which then creates selection pressure for genetic variants that stabilise the adaptive response — without invoking Lamarckian inheritance of acquired characters.
- Independently proposed in 1896 by James Mark Baldwin, C. Lloyd Morgan, and Henry Fairfield Osborn, the idea was largely neglected during the Modern Synthesis but was revived by computational models, most notably Hinton and Nowlan's 1987 simulation demonstrating that learning can smooth a fitness landscape and accelerate the genetic fixation of adaptive traits.
- The Baldwin effect bridges behavioural ecology and evolutionary genetics by proposing that what organisms do can influence what they become over evolutionary time, with potential examples including the evolution of bird song dialects, tool use traditions, and dietary specialisations in primates.
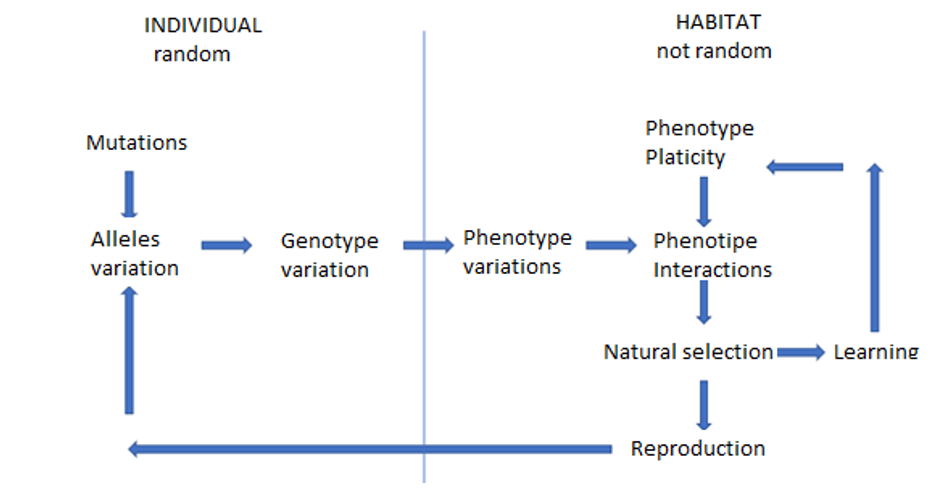
The Baldwin effect is the hypothesis that behaviours learned during an organism's lifetime, or phenotypic changes induced by the environment, can influence the course of genetic evolution without requiring the Lamarckian inheritance of acquired characters. The mechanism proceeds in two stages: first, organisms that can learn or develop adaptive responses to a novel environmental challenge survive and reproduce more successfully than those that cannot, creating selection pressure that favours the capacity for the adaptive response; second, over many generations, natural selection favours genetic variants that produce the adaptive phenotype more reliably, more efficiently, or more constitutively, eventually reducing or eliminating the organism's dependence on learning or environmental induction for its expression. The effect was independently proposed in 1896 by the American psychologist James Mark Baldwin, the British comparative psychologist C. Lloyd Morgan, and the American palaeontologist Henry Fairfield Osborn, each of whom sought to explain how behavioural flexibility could influence evolutionary trajectories within a strictly Darwinian framework.1, 2, 3
{kind=link}
The 1896 proposals
James Mark Baldwin's 1896 paper "A new factor in evolution" proposed that the ability to learn adaptive behaviours could act as a bridge between environmental challenges and genetic change. Baldwin argued that when a population encounters a new environmental demand, individuals that can learn an appropriate behavioural response will survive and reproduce, while those that cannot will be eliminated. This differential survival creates a selection pressure that had not previously existed, and over subsequent generations, genetic variants that predispose organisms toward the adaptive behaviour, or that produce it more efficiently or with less need for learning, will be favoured. The end result is that a trait that was initially acquired through learning becomes progressively more "instinctive" in the population, even though no learned character was directly inherited.1
C. Lloyd Morgan published his formulation in the same year, using the term "organic selection" to describe the process by which individually acquired modifications could shield organisms from selective elimination long enough for coincident heritable variations to accumulate. Morgan was careful to distinguish his proposal from Lamarckism: the learned behaviour itself was not transmitted to offspring, but the capacity for learning, and any genetic variants that happened to produce a similar phenotypic effect, were subject to standard Darwinian selection.2 Henry Fairfield Osborn, also writing in 1896, arrived at a similar conclusion from palaeontological considerations, arguing that behavioural adaptability could explain how organisms appeared to evolve in the direction of their learned responses without invoking the inheritance of acquired characters.3
The convergence of three independent proposals in a single year reflected a broader concern in late nineteenth-century biology about the mechanism of adaptive evolution. Lamarckian inheritance, the idea that organisms could pass on characteristics acquired during their lifetimes, had been increasingly challenged by experimental work, most notably by August Weismann's doctrine of the separation of germ and somatic cells. Baldwin, Morgan, and Osborn each sought to show that the apparent directedness of adaptive evolution could be explained by natural selection acting on heritable variation in behavioural flexibility, without recourse to Lamarckism.1, 2, 7
Relationship to genetic assimilation
The Baldwin effect is closely related to, but not identical with, the process of genetic assimilation described by Conrad Hal Waddington. In Waddington's classic experiments in the 1940s and 1950s, Drosophila melanogaster were subjected to environmental stress (heat shock or ether exposure), which induced a novel phenotype (crossveinless wings or bithorax-like transformations) in a fraction of individuals. By selectively breeding the individuals that showed the induced phenotype, Waddington demonstrated that after several generations the phenotype appeared even without the environmental stimulus, indicating that an initially environmentally triggered response had become genetically constitutive.5, 6
Crispo clarified the relationship between the Baldwin effect and genetic assimilation in a 2007 review. Both processes describe how an initially environmentally contingent phenotype becomes genetically fixed over evolutionary time. The Baldwin effect, as originally formulated, emphasises learned behaviours and behavioural plasticity as the initial adaptive response, while Waddington's genetic assimilation is framed more broadly in terms of developmental plasticity and the canalisation of development. In both cases, the mechanism is orthodox natural selection acting on pre-existing genetic variation that affects the threshold, reliability, or constitutiveness of the phenotypic response. Neither process requires any non-Darwinian mechanism; the apparent Lamarckian flavour arises because the phenotype that is ultimately genetically fixed resembles the one that was initially acquired through environmental induction or learning.7
George Gaylord Simpson discussed the Baldwin effect in his contribution to the 1960 Darwin centennial volume, largely dismissing it as theoretically plausible but empirically unimportant. Simpson's scepticism reflected the broader stance of the Modern Synthesis, which treated behaviour primarily as a product of genetic evolution rather than as a driver of it. This dismissive attitude contributed to the neglect of the Baldwin effect for much of the twentieth century, a situation that only began to change with the advent of computational models and the renewed interest in developmental plasticity in the 1980s and 1990s.16
Computational models
The Baldwin effect was dramatically revived by a computational model published by Geoffrey Hinton and Steven Nowlan in 1987. Their simulation considered an organism with a genotype consisting of 20 binary "genes," each of which could be set to 1, 0, or "?" (learnable). Only the single genotype with all 20 positions set to 1 was fit; all other combinations were equally unfit. Without learning, finding this optimal genotype by random mutation was effectively impossible because the fitness landscape was entirely flat except for a single spike. However, when organisms with "?" alleles were allowed to discover the correct setting through random learning trials during their lifetime, the fitness landscape was smoothed: organisms with more correct alleles fixed in their genotype had a higher probability of discovering the complete solution through learning, and therefore achieved higher average fitness.4
The result was striking. With learning enabled, the population rapidly evolved toward the optimal genotype, with the "?" (learnable) positions being gradually replaced by genetically fixed 1 alleles over generations. Learning effectively converted a fitness landscape with no gradient into one with a gradient that natural selection could climb. Hinton and Nowlan concluded that "the ability to learn can guide the course of evolution," demonstrating in a formal model what Baldwin had argued verbally a century earlier. The simulation provided a clear, mechanistic demonstration that learning could accelerate genetic evolution by smoothing the fitness landscape and creating selection pressure for the genetic fixation of learned traits.4
Subsequent computational studies have explored the conditions under which the Baldwin effect is most effective. The effect is strongest when the fitness landscape is rugged (containing isolated optima that are difficult to find by mutation alone), when the cost of learning is moderate (so that there is selection pressure to replace learning with genetic fixation), and when the environmental challenge is sufficiently stable that genetically fixing the trait provides a reliable advantage over learning it anew each generation. When the environment changes rapidly, plasticity and learning retain their advantage, and genetic fixation does not occur, a prediction that accords with the theoretical expectation that plasticity is favoured in variable environments and constitutive expression is favoured in stable ones.4, 10
The mechanism in detail
The Baldwin effect operates through a sequence of steps that are individually uncontroversial within standard evolutionary theory but combine to produce a pattern that superficially resembles Lamarckian inheritance. First, a population encounters a new environmental challenge, such as a novel food source, predator, or climatic condition. Second, some individuals cope with the challenge through behavioural learning, developmental plasticity, or physiological acclimation, while others cannot and are eliminated by selection. This differential survival constitutes selection for the capacity to produce the adaptive response, whether through learning or plasticity.1, 7
Third, once the population has been stabilised by the learned or plastic response, there is ongoing selection pressure for genetic variants that produce the adaptive phenotype more efficiently, more reliably, or at lower cost. An individual that produces the correct response innately, without the time, energy, and risk involved in learning it, has a fitness advantage over one that must learn it each generation. Over time, alleles that reduce the developmental or learning threshold for producing the adaptive phenotype increase in frequency, and the trait becomes progressively more constitutive and less dependent on environmental induction.7, 8
The critical distinction from Lamarckism is that the learned behaviour or plastic response is never directly transmitted to offspring. What is inherited is genetic variation affecting the propensity, efficiency, or threshold of the adaptive response, and this variation is shaped by natural selection in the standard Darwinian fashion. The Baldwin effect thus provides a mechanism by which organisms' behaviours can shape their selective environments, which in turn shape their genetic evolution, creating a feedback loop between behaviour and genes that operates entirely within the framework of population genetics.1, 7, 9
Proposed examples
Identifying clear empirical examples of the Baldwin effect is challenging because the process operates over evolutionary timescales and involves the replacement of an initially learned or plastic response by a genetically constitutive one, a transition that erases the evidence of its own occurrence once complete. Nevertheless, several proposed examples illustrate the plausibility of the mechanism.
Bird song dialects provide a suggestive case. In many songbird species, males learn their songs from adult tutors during a sensitive period in early development, and the resulting dialects can persist for generations within local populations. Slater reviewed evidence that song learning creates selection pressures on both the neural machinery for learning and the genetic predisposition to learn particular song elements. In species where song functions in mate choice, females may preferentially mate with males that produce local dialect songs, creating sexual selection on the capacity to learn and reproduce the locally adaptive song type. If certain genetic variants predispose males toward producing elements of the favoured dialect more easily, these alleles would be favoured by selection, potentially leading to the genetic canalisation of song structure over evolutionary time.11
Primate tool use offers another candidate. Whiten and colleagues documented extensive cultural traditions in wild chimpanzee populations, including tool-use behaviours such as termite fishing, nut cracking, and ant dipping that are socially learned and vary across communities in ways that cannot be explained by genetic or ecological differences alone.19 If tool use allows chimpanzees to exploit food resources more efficiently, creating selection pressure for cognitive and manual traits that facilitate tool learning, then the Baldwin effect predicts that genetically based improvements in fine motor control, hand morphology, or cognitive flexibility should evolve in populations where tool-use traditions are sustained over long periods. The challenge is distinguishing this prediction from the alternative hypothesis that genetic evolution of cognitive traits drove the emergence of tool use, rather than the reverse.12, 20
Dietary adaptations in human evolution may also involve Baldwin-like dynamics. The adoption of dairying by Neolithic pastoralist populations exposed these groups to a novel dietary resource (fresh milk), and individuals who could tolerate lactose as adults had a nutritional advantage. The subsequent evolution of lactase persistence alleles in multiple pastoralist populations independently suggests that the culturally learned behaviour of dairying created a selective environment that drove genetic change, a sequence consistent with the Baldwin effect operating through cultural rather than individual learning.9, 12
Relationship to niche construction
The Baldwin effect shares conceptual territory with niche construction, the process by which organisms modify their own selective environments through their activities, metabolism, and choices. Odling-Smee, Laland, and Feldman argued that niche construction is a pervasive evolutionary process in which organisms do not merely respond to their environments through adaptation but actively shape the selection pressures that act on themselves and their descendants. Dam building by beavers, soil modification by earthworms, and cultural practices by humans all alter the selective environment in ways that feed back into genetic evolution.13
The Baldwin effect can be viewed as a specific case of niche construction in which the niche modification is behavioural or developmental rather than physical. When an organism learns a new behaviour or develops a plastic response to a novel challenge, it effectively constructs a new selective niche for itself and its descendants, creating selection pressures that would not have existed without the behavioural innovation. The convergence of the Baldwin effect, niche construction, and genetic assimilation under the umbrella of the "extended evolutionary synthesis" reflects a growing recognition that the relationship between organisms and their environments is reciprocal, with behaviour, development, and ecology all feeding back into genetic evolution.13, 14
Controversies and current status
The Baldwin effect has been both celebrated as a neglected insight and criticised as theoretically redundant. Proponents argue that it provides a crucial bridge between behavioural ecology and evolutionary genetics, explaining how organisms' behavioural flexibility can accelerate adaptation, guide populations toward adaptive peaks that would be inaccessible through genetic mutation alone, and initiate evolutionary trajectories that would not otherwise occur. The Hinton and Nowlan simulation provided compelling theoretical support for this view, and the subsequent incorporation of the Baldwin effect into the extended evolutionary synthesis has given it renewed prominence.4, 14, 15
Critics counter that the Baldwin effect is simply natural selection acting on genetic variation in the capacity for learning or plasticity, and that labelling it as a distinct "effect" adds terminology without adding explanatory power beyond standard population genetics. Simpson's 1960 critique set the tone for this view, and some modern population geneticists argue that the conditions under which the Baldwin effect produces qualitatively different outcomes from ordinary directional selection are narrow and rarely met in nature.16
The current status of the Baldwin effect in evolutionary biology reflects a broader debate about the adequacy of the Modern Synthesis. Laland and colleagues have argued that phenomena such as the Baldwin effect, niche construction, and developmental plasticity require an extension of evolutionary theory beyond the gene-centred framework of the Modern Synthesis, while others maintain that these phenomena are already accommodated within standard evolutionary theory and do not require any conceptual revision. Regardless of where one stands in this debate, the Baldwin effect has contributed a valuable perspective: that what organisms do during their lifetimes is not merely a consequence of their evolutionary history but can actively shape their evolutionary future.14, 15, 17
References
A mode of evolution requiring neither natural selection nor the inheritance of acquired characters
The Baldwin effect and genetic assimilation: revisiting two mechanisms of evolutionary change mediated by phenotypic plasticity
Evolution in four dimensions: genetic, epigenetic, behavioral, and symbolic variation in the history of life
The Baldwin effect: a neglected influence on C. H. Waddington's use of genetic assimilation