Overview
- Cope's rule is the generalisation that lineages of animals tend to evolve toward larger body size over geological time, an observation associated with the 19th-century American palaeontologist Edward Drinker Cope but actually formulated by Bernhard Rensch in 1948 from work by Charles Depéret.
- The rule has been confirmed quantitatively in some clades, including North American Cenozoic mammals (where new species average 9.1% larger than their ancestors), Mesozoic dinosaurs, marine animals (a 150-fold increase in mean biovolume since the Cambrian), and Paleozoic brachiopods, but rejected in others such as insects, birds, Cretaceous bivalves, and modern oryzomyine rodents.
- The central debate is whether the pattern reflects a driven trend of within-lineage selection for larger size or a passive diffusion away from a lower size limit imposed by physiology, with current evidence pointing to a mixture of both depending on clade and timescale.
Cope's rule is the generalisation that lineages of animals tend, over geological time, to evolve toward larger body size. The pattern is named for the 19th-century American palaeontologist Edward Drinker Cope, although Cope himself never explicitly formulated it; the modern statement of the rule was crystallised by Bernhard Rensch in 1948, building on a 1907 chapter by the French palaeontologist Charles Depéret entitled “the law of increase in size in phyletic branches.”5 The rule occupies an unusual position in evolutionary biology: it is one of the most frequently invoked macroevolutionary generalisations, repeatedly demonstrated in some clades and emphatically rejected in others, and the mechanism behind it has been disputed for more than half a century.
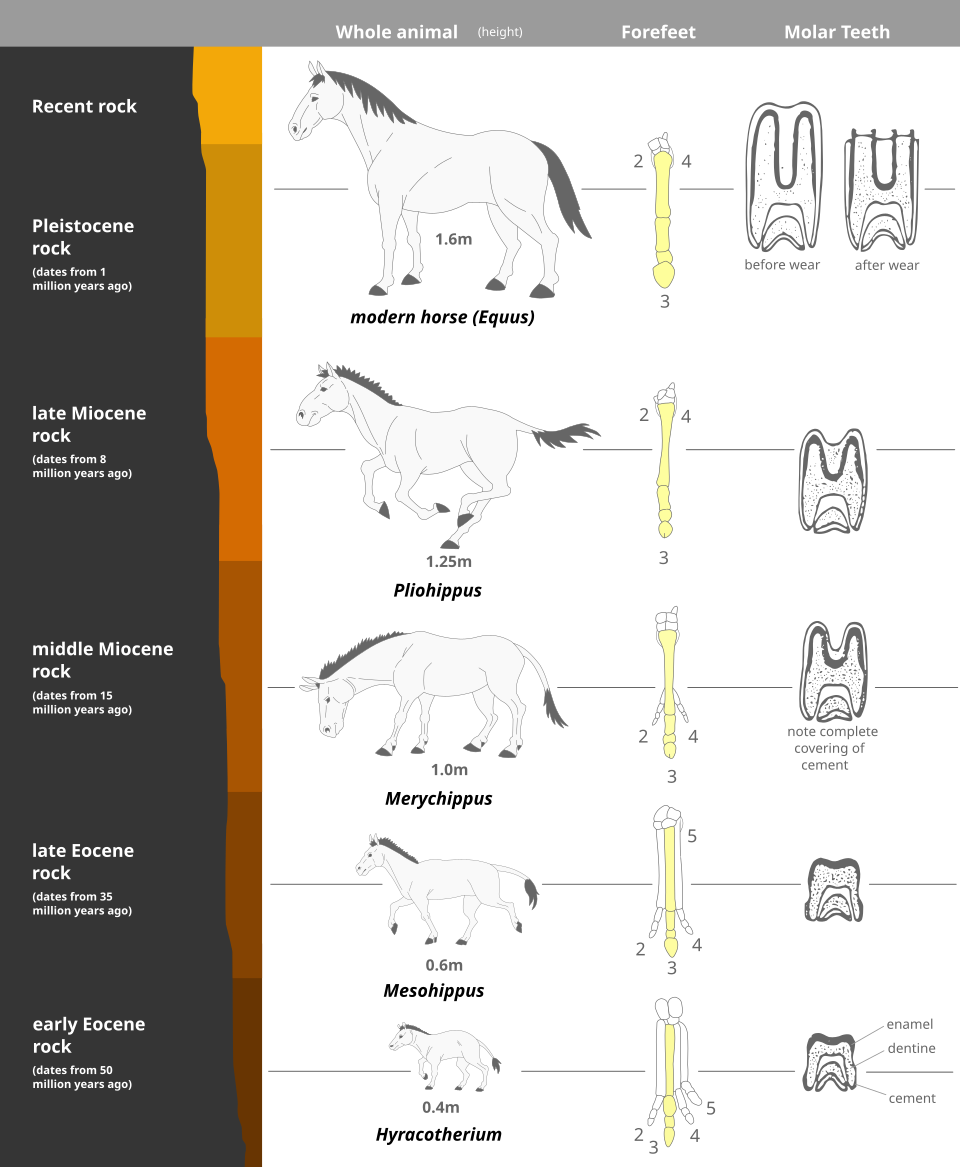
The intuitive appeal of Cope's rule is grounded in the visible parade of fossil giants. The earliest known horse, Hyracotherium, stood roughly 40 centimetres at the shoulder in the Eocene; the modern Equus stands more than 1.6 metres. The earliest sauropodomorph dinosaurs of the Triassic weighed a few tens of kilograms; by the Cretaceous, Argentinosaurus may have weighed nearly 100 tonnes.12, 19 Yet what looks like a universal march toward gigantism dissolves on closer inspection. Many lineages remain small, others shrink, and the question of whether the apparent trend reflects an active evolutionary force or a statistical artefact of where life began — near the lower physiological limit of body size — lies at the heart of the modern debate over Cope's rule.4, 6, 8
Origins of the rule and the misattribution to Cope
Edward Drinker Cope (1840–1897) was one of the most prolific vertebrate palaeontologists of the 19th century. A wealthy Quaker autodidact who described more than a thousand species of fossil vertebrates and published roughly six hundred scientific papers over his career, Cope is best known today for his ferocious rivalry with Othniel Charles Marsh during the so-called Bone Wars, a period of cutthroat dinosaur prospecting in the American West that ran from the late 1870s into the early 1890s.1 In his collection of essays The Origin of the Fittest (1887), Cope advanced an explicitly orthogenetic view of evolution, arguing that lineages tended to follow predetermined trajectories toward greater specialisation, larger size, and more complex morphology, and that these trajectories were driven by an internal “law of acceleration” rather than by natural selection alone.1

{kind=link}
The narrower claim that body size tends to increase within lineages does not appear in Cope's writings in the form now associated with his name. The historical attribution was traced by David Hone and Michael Benton, who showed that the modern rule was first stated as “Cope's rule” by the German evolutionary biologist Bernhard Rensch in 1948, and that Rensch's proximate source was actually Charles Depéret's 1907 monograph Les transformations du monde animal, which contained an entire chapter titled “the law of increase in size in phyletic branches.”5 Cope's general orthogenetic framework was sufficiently consistent with Depéret's claim that later authors attached his name to it without close attention to the original texts. The misattribution is now firmly entrenched in the literature, and the term Cope's rule is universally used despite the more accurate label “Depéret's rule” that Hone, Benton, and others have occasionally tried to popularise.5
Independent of nomenclature, the empirical observation that many fossil lineages contain progressively larger species was already familiar to 19th-century palaeontologists. Othniel Marsh's reconstruction of the horse lineage from Eocene Hyracotherium through Mesohippus, Merychippus and Pliohippus to modern Equus was promoted as a paradigmatic example of progressive body-size increase as early as the 1870s, and remained the textbook illustration of macroevolutionary directionality for the better part of a century.20 Modern phylogenetic analyses have substantially complicated that picture, but the visual force of the horse series was instrumental in establishing the intuition that Rensch later codified.
Alroy and the Cenozoic mammal test
The first quantitatively rigorous test of Cope's rule on a continental scale was published by John Alroy in Science in 1998. Alroy compiled body-mass estimates for 1,534 species of North American Cenozoic mammals and, crucially, used genus membership to identify ancestor-descendant pairs within lineages, allowing him to ask whether new species really were larger than the older species in the same genus.2 The answer was unambiguous: across the entire 65-million-year Cenozoic record, new species averaged 9.1 percent larger in body mass than the older congeneric species they replaced. The effect persisted throughout the Cenozoic, was not attributable to sampling bias, and was strongest among larger mammals, dropping nearly to zero for species below about one kilogram.2
Alroy's design was important because it bypassed the central methodological objection to most earlier tests of Cope's rule. Earlier studies had compared the average body size of an entire clade at successive time intervals, but a clade's mean body size can drift upward simply because new lineages of large body size are added to the clade through immigration or origination, even if no individual lineage is increasing in size at all. By restricting attention to within-genus successions, Alroy isolated the within-lineage component of size change and showed that, for North American Cenozoic mammals at least, it really was directional.2 The result has since been replicated and extended. Felisa Smith and colleagues, working on the maximum body size of terrestrial mammals on each continent, found a near-exponential increase in maximum mass after the Cretaceous-Paleogene boundary, with maxima levelling off only around 40 million years ago and remaining roughly constant thereafter.10
Within-lineage body-size change across major animal groups2, 7, 13, 14
Subsequent work has refined the mammal picture in two important ways. First, the within-lineage signal is not uniform: it is concentrated in particular ecomorphological guilds, with herbivores and large carnivores showing stronger trends than small insectivores and frugivores.18 Second, when extant mammal lineages alone are analysed without their fossil ancestors, the trend largely disappears, suggesting that the directional signal recovered from the deep Cenozoic record is partly an artefact of survivorship: the small-bodied lineages that did not increase in size also did not leave many descendants in the modern fauna.15
The marine record
If Cope's rule is to be tested at the largest possible taxonomic scale, the marine fossil record offers the most complete dataset. Noel Heim and colleagues, working at Stanford and Swarthmore, compiled body-size estimates for 17,208 genera of marine animals spanning the past 542 million years and asked whether mean, minimum, and maximum body size had changed systematically since the Cambrian explosion.7 Their result was striking: mean biovolume across genera increased by a factor of 150, while minimum biovolume decreased by less than a factor of ten and maximum biovolume increased by more than a factor of 100,000. The increase in mean size was substantially greater than would be predicted by neutral diffusion from a Cambrian starting distribution and was driven primarily by differential diversification — classes with larger mean body size, such as vertebrates and large arthropods, diversified faster than classes with smaller mean body size.7
The Heim study marked an important shift in how Cope's rule is conceptualised. Where Alroy had focused on within-lineage anagenesis, Heim and colleagues showed that even at the level of entire phyla and classes, an apparent directional trend can arise without any individual lineage being under selection for larger size. The mechanism in their analysis was differential origination and extinction: classes happened to be founded at different mean sizes, and those with larger founding sizes happened to diversify more rapidly. The pattern is real, but it is not anagenetic; it is a sorting process operating at the level of higher taxa, similar in spirit to the species-selection arguments developed by Stephen Jay Gould and Steven Stanley in the 1970s.7
The marine bivalve record presents a sharp contrast and motivated one of the most influential critiques of Cope's rule. David Jablonski's 1997 census of body-size changes in 191 lineages of Late Cretaceous molluscs found that net within-lineage size increases were no more common than net decreases, and that most lineages showed no net trend at all.3 The clade-level signal of increasing maximum size in Cretaceous bivalves arose almost entirely from an expansion of the clade's size range — the addition of a few large outlier species — rather than from any tendency of individual lineages to grow larger over time. Jablonski concluded that previously claimed examples of Cope's rule should be re-examined to distinguish phyletic anagenesis from passive expansion of the size envelope, and his paper effectively reset the methodological standards for the field.3
Dinosaurs and the rise of the giants
Dinosaurs are the standard case study in Cope's rule because the body-size range of the clade spans nearly eight orders of magnitude, from the smallest known maniraptorans of perhaps 30 grams to titanosaurs estimated at upwards of 70,000 kilograms.12 Roger Benson and colleagues used non-uniform Ornstein-Uhlenbeck and trend models to fit body-size evolution across more than 400 dinosaur species and found that dinosaur body-size evolution is best described as movement on a multimodal adaptive landscape, with discrete optima that occasionally undergo abrupt shifts.12 Within most dinosaur subclades, including theropods and ornithischians, an active Cope's-rule trend toward larger size is detectable. Cretaceous sauropods, however, are an exception: after their initial Triassic-Jurassic gigantism, their body-size distribution remained roughly stable for the remainder of the Mesozoic, suggesting that they had reached an upper limit imposed by physiology, locomotor mechanics, or ecology.12
The earliest stages of dinosaur body-size evolution are also informative. Apaldetti and colleagues described a Late Triassic sauropodomorph from Argentina, Ingentia prima, with a body mass of roughly 10 tonnes, demonstrating that gigantism in this lineage was achieved at least 30 million years earlier than was previously thought and that the giant body plan evolved independently of the column-like limbs and pneumatised skeletons characteristic of later sauropods.19 The trend toward sauropod gigantism was not, in this view, a slow accumulation of incremental size increases but a relatively rapid early jump.
The most comprehensive challenge to a simple Cope's-rule reading of dinosaur evolution comes from work on the broader archosauromorph radiation that preceded the dinosaurs. Sookias, Butler, and Benson analysed body-size data for more than 400 species of therapsids (mammal ancestors) and archosauromorphs (including dinosaurs) spanning the Late Permian through the Middle Jurassic, and found that maximum-likelihood model fitting strongly favoured passive trait evolution over active Cope's-rule processes in most clades.11 Their conclusion was that the conspicuous body-size turnover associated with the rise of dinosaurs — the replacement of large-bodied therapsids by even larger-bodied archosauromorphs — was driven by passive diffusion of trait values away from a small-bodied lower bound, not by active selection for larger size operating within lineages.11
Driven trends, passive trends, and the left wall
The conceptual framework for distinguishing kinds of macroevolutionary trends was developed by Daniel McShea in 1994 in a paper that has become foundational to the modern Cope's-rule literature.6 McShea distinguished driven trends, in which the state variable (body size, in this case) tends to change in the same direction in most lineages, from passive trends, in which lineages diffuse roughly randomly in trait space but are constrained on one side by a hard lower limit. A passive trend produces an apparent increase in mean and maximum body size simply because there is more room to grow than to shrink.6
The distinction matters because the two kinds of trend have radically different evolutionary implications. A driven trend implies that selection consistently favours larger body size in most lineages most of the time, which in turn implies that there is some general fitness advantage to being larger. A passive trend implies no such thing: the appearance of directional evolution emerges from the geometry of the size distribution alone, with no need for selection to favour any particular direction. McShea proposed several empirical tests for distinguishing the two, the most widely used of which is the minimum-tracking test: if the minimum body size in a clade increases through time alongside the mean and maximum, the trend is driven; if the minimum stays flat while the mean and maximum increase, the trend is passive.6
Driven versus passive trends: predictions and tests6, 7, 11
| Property | Driven trend | Passive trend |
|---|---|---|
| Mean body size over time | Increases | Increases |
| Maximum body size over time | Increases | Increases |
| Minimum body size over time | Increases | Roughly constant |
| Skewness of size distribution | Symmetric or shifting | Right-skewed, anchored at left wall |
| Cause | Within-lineage selection for larger size | Diffusion bounded by physiological lower limit |
| Example | Cenozoic mammals (Alroy 1998) | Triassic archosauromorphs (Sookias 2012) |
Stephen Jay Gould articulated the strongest version of the passive-trend critique in a 1997 commentary in Nature entitled “Cope's rule as psychological artefact,” written in response to Jablonski's paper in the same issue.8 Gould argued that the apparent directionality of life's history toward larger size, greater complexity, and more elaborate morphology was an inevitable consequence of starting from the absolute lower bound of what was physically possible. Bacteria mark the left wall of life's body-size distribution, and any subsequent diffusion can only move outward into previously unoccupied size classes. The increasing maximum size of life over the past 3.5 billion years, in this view, requires no special explanation: it is what an unconstrained random walk from a left wall must produce.8 Gould's argument did not deny that some lineages do exhibit driven Cope's-rule trends, but it insisted that the burden of proof should rest on demonstrating directional selection rather than on assuming it.
Stanley had anticipated parts of this argument in his 1973 paper on Cope's rule, in which he proposed that smaller-bodied species are more likely to speciate than larger-bodied species because they tend to have larger populations, shorter generation times, and a greater capacity to form peripheral isolates.4 Under this model, new lineages preferentially originate from small ancestors, which then have nowhere to go but up; the apparent directional trend is a consequence of asymmetric speciation rather than asymmetric within-lineage selection. Stanley's mechanism is a kind of species-level passive trend, distinct from McShea's diffusion model but predictively similar at the clade level.4
Adaptive explanations for size increase
The case for a driven Cope's-rule trend rests on the observation that, in many ecological contexts, larger body size carries demonstrable fitness advantages. Maurer, Brown, and Rusler argued in 1986 that body size correlates positively with ecological dominance — the ability to monopolise food resources, defend territories, and resist predation — and that selection for ecological dominance could plausibly produce a driven trend in many vertebrate clades.23 Subsequent reviews by Hone and Benton catalogued the specific advantages associated with larger size: improved thermoregulation through reduced surface-to-volume ratios, expanded prey range for predators, reduced vulnerability to predation for prey, increased fecundity and offspring survival, greater starvation resistance, and improved competitive ability in intraspecific contests.5
The pinniped record provides a particularly clean example of an apparently driven Cope's-rule trend in a marine clade. Churchill, Clementz, and Kohno reconstructed the body-size history of the Pinnipedimorpha (seals, sea lions, and walruses) and found a near-monotonic increase in mean body size from the Oligocene origin of the group through the Pleistocene, with the largest pinnipeds (modern elephant seals and the extinct desmostylians) appearing in the late Cenozoic.24 The pinniped trend passes the minimum-tracking test — minimum body size increases alongside the mean — and is consistent with selection for thermoregulatory efficiency in cold marine environments and for competitive success in male-male combat over breeding territories.24
A more recent and ambitious framework has been proposed by Jonathan Saulsbury and colleagues, who argued that the prevalence of Cope's-rule trends in any given clade is determined by ecological context rather than by intrinsic clade properties.25 Drawing on a comparative analysis across multiple invertebrate groups, they showed that Cope's rule predominates in clades occupying productive, predator-rich environments where large body size confers escape from predation and access to a broader prey spectrum, while the inverse pattern (decreasing size over time) predominates in resource-limited environments where small size confers metabolic efficiency and shorter generation times. The implication is that Cope's rule is not a single phenomenon but an outcome that can be predicted from ecological first principles.25
Exceptions and the inverse rule
Cope's rule is not universal, and the exceptions are as illuminating as the confirmations. Insects represent the most conspicuous failure of the rule among major animal groups. Waller and Svensson examined body-size evolution in odonates (dragonflies and damselflies), an old insect order with a continuous fossil record extending more than 250 million years, and found no detectable Cope's-rule trend despite consistent positive selection on adult body size in modern populations.14 They proposed that the absence of a long-term size increase reflects a tradeoff between the fitness benefits of larger adult size and the developmental costs of longer larval periods, which expose juveniles to higher predation and starvation risk. Insects more broadly are constrained by the physics of tracheal oxygen delivery, which sets an upper limit on body size that has not been substantially exceeded since the Carboniferous, when atmospheric oxygen concentrations were higher.
Modern oryzomyine rodents likewise show no Cope's-rule pattern. Maestri and colleagues reconstructed body-mass evolution in this widespread Neotropical group and found a tendency toward miniaturisation, not gigantism, across the past several million years.15 The result is consistent with selection for shorter generation times and higher reproductive output in the rapidly fluctuating environments of South American grasslands and may reflect a general pattern in small mammals that contrasts sharply with the trend Alroy documented for larger Cenozoic mammals.15
The most dramatic exceptions, however, are the cases of insular dwarfism. When large mammals colonise islands, they consistently evolve smaller body sizes, often within a few thousand generations. Pleistocene dwarf elephants on Mediterranean islands such as Crete, Sicily, Malta, and Cyprus reached body masses as small as 200 kilograms, roughly two percent of mainland straight-tusked elephant mass; a single Cretan dwarf mammoth, Mammuthus creticus, may have weighed only about 300 kilograms, the smallest mammoth known.17 Mark Lomolino's synthesis of insular size evolution showed that the direction of size change on islands depends predictably on starting size: small species become larger (insular gigantism) and large species become smaller (insular dwarfism), converging toward an intermediate optimum in the absence of large predators and with limited resources.16 Insular dwarfism is not a contradiction of Cope's rule so much as an inversion of it driven by a different ecological context, and it lends support to Saulsbury and colleagues' argument that the direction of body-size evolution is contingent on ecological circumstance.25
The extinction cost of being big
Even when a Cope's-rule trend is real and driven by selection, the long-term consequences for the lineages that follow it can be lethal. Blaire Van Valkenburgh, Xiaoming Wang, and John Damuth documented this pattern with unusual clarity in two extinct subfamilies of North American canids, the Hesperocyoninae and Borophaginae.9 Both groups exhibited textbook Cope's-rule trends over their multi-million-year histories, with successive species becoming progressively larger and more carnivorous. In each case, the largest, most hypercarnivorous species had the shortest mean species durations and accounted for the bulk of extinctions; the smaller, more generalist species in both subfamilies persisted longer.9 The same pattern of increasing size leading to dietary specialisation, increased vulnerability to environmental perturbation, and elevated extinction risk has since been documented in saber-toothed cats, marine reptiles, and several Cenozoic ungulate radiations.
Van Valkenburgh and colleagues coined the phrase “the macroevolutionary ratchet” for this dynamic: selection for individual fitness drives lineages toward larger size and greater specialisation, but the resulting populations are smaller, more energetically demanding, and less resilient to ecological disruption, so the same trend that benefits individuals undermines the persistence of clades.9 The macroevolutionary ratchet helps explain a puzzle that has long bothered palaeontologists: why so many of the giant land mammals of the late Cenozoic — mammoths, mastodons, ground sloths, glyptodonts, woolly rhinoceroses, the giant short-faced bear — vanished during the climatic and human-induced disturbances of the Pleistocene-Holocene transition, while smaller species in the same orders persist today. Cope's rule, at the timescale of individual lineages, may be a slow march toward the cliff edge.9
{kind=link}
{kind=link}
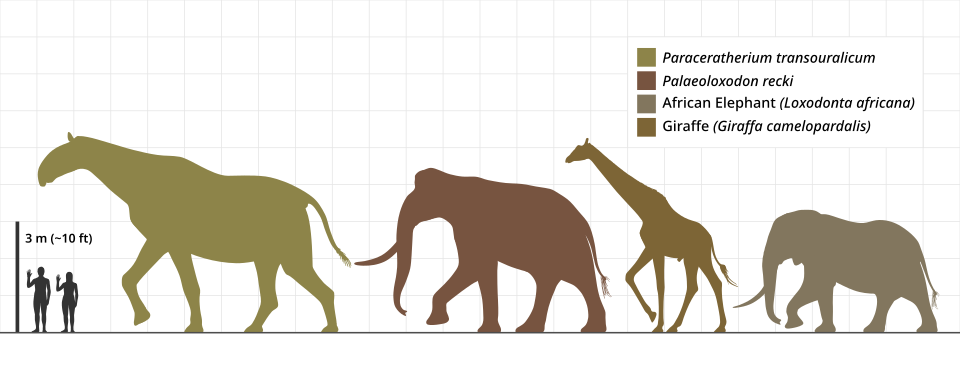
The largest land mammal that ever lived, Paraceratherium transouralicum, exemplifies the Van Valkenburgh dynamic. This Oligocene hornless rhinocerotoid stood approximately 4.8 metres at the shoulder; Larramendi's volumetric reconstructions place its body mass at roughly 17 tonnes, comparable to or slightly exceeding the mass of three or four modern African elephants, and the genus ranged across central Asia from what is now Pakistan to Mongolia.22 The lineage persisted for only about 11 million years before disappearing in the late Oligocene, probably as a result of climate-driven shrinking of the open woodland habitats it required. Paraceratherium illustrates the same pattern Van Valkenburgh and colleagues documented in canids: a clade pursues larger body size, achieves a maximum that strains the physiological and ecological limits of what is possible on land, and then disappears.9, 22
Tests in brachiopods and foraminifera
Two further studies have shaped the modern understanding of how Cope's rule operates at different taxonomic scales. Phillip Novack-Gottshall and Michael Lanier examined body-size evolution in Paleozoic brachiopods over a 250-million-year interval and found that net increases in mean body size at the class level arose not from within-lineage selection in constituent families but from the preferential origination of new families at relatively large body sizes.13 Within individual brachiopod families, Cope's-rule trends were rare; the class-level signal was a sorting effect operating across families. Their conclusion was that the apparent universality of Cope's rule depends sensitively on the taxonomic scale at which the test is conducted: a clade can show a strong directional trend in mean body size at high taxonomic levels and no trend at all within its component lineages.13
Anthony Arnold, Christopher Kelly, and William Parker conducted an analogous test in planktonic foraminifera, a group whose continuous deep-sea sediment record makes them ideal for high-resolution studies of body-size change.21 They found that Cenozoic planktonic foraminifera do exhibit a net increase in mean test size, but that the increase is concentrated in the early diversification of new lineages rather than in within-lineage anagenesis. Once the data are corrected for size-related preservation biases and for the effects of new lineage origination at small body sizes, the within-lineage signal is much weaker than initial inspection suggests. Arnold and colleagues argued that smaller-bodied foraminifera species are more adaptively responsive to environmental change because of their shorter generation times, and that the apparent Cope's-rule pattern is driven by the differential survivorship of large-bodied descendants of these short-generation-time founders.21
These studies, together with Heim and colleagues' analysis of marine animals as a whole, have produced a consensus picture in which Cope's-rule patterns can arise through at least four distinct mechanisms: within-lineage selection for larger size (true anagenetic Cope's rule); differential origination of new lineages at large body sizes (lineage-sorting); differential extinction of small-bodied lineages (filtering); and passive diffusion away from a left wall (Gould's bacterial argument). Disentangling these mechanisms requires the kind of fine-grained ancestor-descendant data that are available only in well-sampled clades, which is one reason why the rule has been confirmed most clearly in groups with exceptional fossil records and rejected in groups whose phylogenies must be inferred from molecular data alone.
Contemporary status of the rule
After more than a century of debate and several decades of quantitative testing, the status of Cope's rule can be summarised in three propositions. First, the rule is real in the limited sense that, among well-studied animal clades with good fossil records, a substantial majority show some form of body-size increase over geological time, whether at the level of within-lineage anagenesis, lineage sorting, or passive diffusion.2, 7, 10, 12, 24 Second, the rule is not a universal law: insects, modern small mammals, Cretaceous bivalves, and many marine invertebrate groups show no Cope's-rule signal, and insular populations consistently violate it.3, 14, 15, 16 Third, the underlying mechanism is heterogeneous: in some clades the trend is genuinely driven by within-lineage selection, in others it reflects sorting among lineages of different mean size, and in still others it is a passive consequence of starting near a lower physiological bound.6, 8, 11
The most productive recent work has moved away from asking whether Cope's rule is true and toward asking under what ecological and developmental conditions body size tends to increase in a given clade. Saulsbury and colleagues' 2023 analysis of the ecological determinants of Cope's rule and its inverse argues that the question is no longer whether body size increases over time but which environments select for which direction of change, and the answer depends on the productivity of the habitat, the predator-prey context, and the metabolic constraints of the organism.25 Cope's rule, on this view, is not a general feature of evolution but a particular outcome that emerges when ecological conditions favour larger size, and which can reverse cleanly when those conditions change.
The historical curiosity that Cope himself never formulated the rule that bears his name is perhaps fitting. The intuition the rule captures — that life as a whole has tended toward larger and more imposing forms — is older than evolutionary biology and may owe more to the visual experience of walking through a fossil hall than to any general macroevolutionary principle. The achievement of the past five decades has been to convert that intuition into a set of empirically testable claims, and the result of the testing is that the intuition is correct in some clades, wrong in others, and almost always more complicated than its 19th-century proponents imagined. Cope's rule, properly understood, is less a law than a starting point for asking better questions about how body size evolves.
References
Rise of dinosaurs reveals major body-size transitions are driven by passive processes of trait evolution
Body size evolution in an old insect order: No evidence for Cope's Rule in spite of fitness benefits of large size