Overview
- Punctuated equilibrium proposes that most species exhibit long periods of morphological stasis lasting millions of years, interrupted by geologically brief episodes of rapid change concentrated during speciation events, a pattern Niles Eldredge and Stephen Jay Gould identified in the fossil record in 1972.
- The theory challenged the gradualist expectation that evolution proceeds by slow, continuous transformation within lineages, instead arguing that the dominant signal in the fossil record is stability rather than directional change, and that most morphological novelty originates in small, peripherally isolated populations during cladogenetic events.
- Decades of quantitative paleontological studies across diverse taxa have confirmed that stasis is a real and pervasive pattern rather than an artifact of an incomplete fossil record, though the relative contributions of punctuated and gradualistic modes vary among lineages and traits.
Punctuated equilibrium is a theory of evolutionary tempo and mode proposing that most species undergo little morphological change for the majority of their geological duration — a condition termed stasis — and that significant evolutionary change is concentrated in geologically brief episodes associated with the branching events by which new species originate. The theory was articulated by the American paleontologists Niles Eldredge and Stephen Jay Gould in 1972 as an explicit challenge to the prevailing assumption that evolution proceeds primarily through the slow, continuous, gradual transformation of entire lineages over time.1 By arguing that the dominant pattern in the fossil record is not gradual directional change but prolonged stability punctuated by rapid divergence, Eldredge and Gould reframed the relationship between paleontology and evolutionary biology and provoked one of the most sustained debates in the history of the life sciences.2, 3
The theory drew on Ernst Mayr's model of peripatric speciation — the origin of new species in small, geographically isolated populations at the margins of the ancestral range — and applied it to the interpretation of fossil sequences. If most evolutionary change occurs rapidly in small peripheral populations that subsequently expand and appear in the fossil record as apparently new forms, then the expectation for any well-sampled fossil lineage should be long intervals of stability interrupted by the abrupt appearance of descendant species, not a smooth continuum of transitional forms.1, 9
Intellectual origins
The intellectual foundations of punctuated equilibrium lie in the mid-twentieth-century Modern Synthesis, particularly in the tension between the way population geneticists and paleontologists understood evolutionary tempo.

{kind=link}
George Gaylord Simpson's landmark 1944 work Tempo and Mode in Evolution had already distinguished several rates of evolution observable in the fossil record, recognizing that some lineages changed slowly and steadily while others appeared to undergo bursts of rapid transformation that he termed quantum evolution.8 Simpson interpreted quantum evolution as occurring when small populations crossed adaptive thresholds, but he embedded this idea within a broadly gradualist framework and did not challenge the assumption that the gaps in the fossil record between species were artifacts of incomplete preservation.
Ernst Mayr's 1942 model of geographic (allopatric) speciation provided the other critical ingredient. Mayr argued that new species arise most commonly when a small population becomes geographically isolated from the parent species and undergoes rapid genetic reorganization in its new environment — a process he later called a genetic revolution.9 Because such peripheral isolates are small and geographically restricted, they are unlikely to be preserved in the fossil record during their brief period of active divergence. Their appearance in the record, if they succeed and expand, would therefore be sudden rather than gradual.
Eldredge's doctoral work on the Devonian trilobite Phacops rana in the 1960s provided the empirical catalyst. Examining sequences of trilobite-bearing limestones across the eastern United States, he found that Phacops populations maintained stable eye morphology (measured by the number of lens columns) for millions of years, with abrupt shifts between distinct morphological states that coincided with the appearance of new species. The pattern was not what gradualism predicted: instead of smooth, continuous change, the record showed prolonged stasis interrupted by sudden transitions.1, 4 Eldredge published his initial findings in 1971, and the following year he and Gould synthesized the broader theoretical framework in their influential chapter "Punctuated equilibria: an alternative to phyletic gradualism."1
Core claims of the theory
Punctuated equilibrium makes two linked empirical claims about the tempo and mode of morphological evolution in sexually reproducing species.
{kind=link}
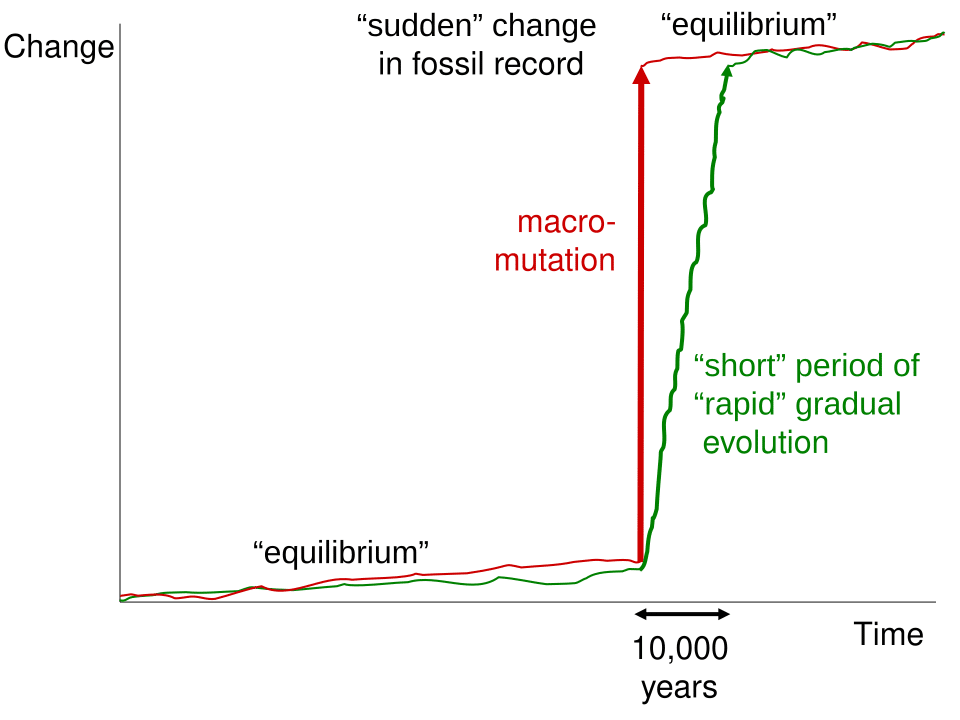
The first is stasis: that species, once established, typically exhibit little net directional morphological change throughout their geological duration, which may span millions to tens of millions of years. The second is punctuation: that most morphological divergence is concentrated in geologically rapid episodes of speciation, during which a descendant species diverges from its ancestor in a time interval that, although it may encompass tens of thousands of years in absolute terms, is too brief to be resolved in most stratigraphic sequences and therefore appears abrupt in the fossil record.1, 2
It is important to note what the theory does not claim. Punctuated equilibrium does not propose that speciation occurs instantaneously or by mechanisms unknown to genetics. The "rapidity" of change is relative to geological timescales: a speciation event lasting 50,000 years would be effectively instantaneous in a stratigraphic section spanning five million years, appearing as a single bedding plane rather than a gradual transition. Nor does the theory deny that natural selection is the primary agent of adaptive change; rather, it proposes that selection within established species tends to maintain existing adaptations (stabilizing selection) rather than driving sustained directional transformation.2, 3
Gould and Eldredge situated their two empirical claims within a broader hierarchical framework of evolutionary theory. If most morphological change is concentrated at speciation events, then the differential origination and extinction of species — a process Gould called species selection — becomes a macroevolutionary force in its own right, analogous to natural selection operating on individuals but at a higher level of the biological hierarchy.3, 15 In this view, long-term evolutionary trends reflect not the gradual transformation of lineages but the differential proliferation of species possessing certain heritable properties. The hierarchical extension proved more controversial than the empirical claims of stasis and punctuation themselves.12, 15
Stasis as a biological phenomenon
Of the two central claims, stasis has attracted the greater volume of empirical investigation and has proved to be the more robust.

_1_(36800705542).jpg){kind=link}
Before Eldredge and Gould drew attention to it, stasis was not considered a phenomenon requiring explanation; it was either ignored or attributed to the imperfection of the fossil record, which was assumed to obscure the gradual changes that must have been occurring. Punctuated equilibrium reframed stasis as a positive result — a real biological signal demanding a causal explanation.1, 6
Quantitative analyses of fossil time series have since confirmed that stasis is a genuine and widespread pattern. Gene Hunt's 2007 study fitted three alternative models — stasis, random walk, and directional change — to 251 fossil morphological time series spanning a range of animal taxa and found that stasis was the best-supported model in the majority of cases, with random walk the next most common and sustained directional change the rarest pattern.5 This result held across a wide range of organisms, environments, and timescales, indicating that the predominance of stasis is not an artifact of any particular taxon or preservational setting.
Among the most compelling case studies are the cheilostome bryozoans studied by Alan Cheetham and Jeremy Jackson. Working with Caribbean fossil species that have exceptionally complete stratigraphic records spanning millions of years, Cheetham and Jackson conducted rigorous morphometric analyses and found that individual species maintained stable multivariate morphologies over intervals of two to six million years, with new species appearing abruptly in the record and coexisting with their morphologically static ancestors.14 The bryozoan data were particularly powerful because the organisms are colonial, clonal, and abundant, minimizing the sampling problems that plague studies of rarer fossils.
Several mechanisms have been proposed to explain why species remain morphologically stable for extended periods. Stabilizing selection — natural selection that favours the existing mean phenotype and eliminates deviants — is the most straightforward, but it requires that the optimal phenotype remain constant despite environmental fluctuations over millions of years. Developmental constraints may limit the range of phenotypic variation available to selection, channelling potential change along a few permitted trajectories and preventing departure from established morphologies. Habitat tracking, in which species shift their geographic ranges to follow preferred environments rather than adapting in place, offers another route to apparent stasis: the species does not change because it moves to maintain its ancestral conditions.6, 13 Finally, gene flow across large, geographically widespread populations may swamp locally adapted variants and prevent any subpopulation from diverging significantly from the species mean, a mechanism that simultaneously explains why speciation tends to occur in peripherally isolated populations where gene flow is disrupted.9, 13
The gradualism debate
The publication of punctuated equilibrium ignited a vigorous and sometimes acrimonious debate that dominated evolutionary paleobiology through the 1980s and 1990s. Critics, particularly population geneticists, raised several objections. Brian Charlesworth, Russell Lande, and Montgomery Slatkin argued in a widely cited 1982 paper that the morphological changes documented in punctuated patterns were fully compatible with standard population-genetic models of gradual change operating over thousands of generations, and that no novel evolutionary mechanisms were required to explain them.17 From this perspective, punctuated equilibrium was a description of a pattern in the fossil record, not a new theory of evolutionary process.
The genetic revolution model that Mayr had proposed as the mechanism driving rapid change in small peripheral populations also came under scrutiny. Nicholas Barton and Brian Charlesworth reviewed the theoretical basis for founder-effect speciation and concluded that genetic drift in small populations was unlikely to produce the kind of coordinated morphological change seen in the fossil record, particularly when opposed by even moderate natural selection.18 The theoretical consensus that emerged was that rapid morphological change in small populations is better explained by strong natural selection in novel environments than by genetic drift or genetic revolutions.
Gould and Eldredge acknowledged some of these criticisms while maintaining that the core empirical observations of stasis and punctuation remained robust regardless of which microevolutionary mechanisms were invoked to explain them. In their 1993 retrospective, they emphasized that the debate had shifted the burden of proof: before punctuated equilibrium, stasis had been dismissed as uninformative absence of data, but it was now recognized as a pattern requiring explanation, and the assumption that all evolution must be gradualistic had been effectively challenged.2 They also clarified that punctuated equilibrium was intended as a claim about the relative frequency of evolutionary modes, not an assertion that gradualism never occurs. Some lineages do exhibit gradual directional change, but such cases are the exception rather than the rule.2, 3
Peter Sheldon's 1987 study of eight trilobite lineages in Ordovician strata of central Wales provided one of the most frequently cited examples of gradualism in the fossil record. Sheldon documented slow, continuous, directional change in pygidial rib counts across all eight lineages over approximately three million years, a pattern more consistent with phyletic gradualism than with punctuated equilibrium.11 Studies of this kind demonstrated that gradualism is a real evolutionary mode, even as the broader literature increasingly confirmed that stasis is more common.
Quantitative evidence
The development of quantitative methods for analysing evolutionary rates in fossil time series transformed the punctuated equilibrium debate from a qualitative argument into a testable empirical question. Hunt's 2006 and 2007 studies were pivotal in this regard. By fitting competing statistical models (unbiased random walk, directional change, and stasis) to hundreds of fossil trait series using maximum-likelihood methods, Hunt showed that the data could discriminate among alternative evolutionary modes with reasonable statistical power.19, 5
Distribution of best-fit evolutionary models across 251 fossil time series5
The results of Hunt's meta-analysis revealed that stasis was the single most common pattern, characterizing approximately 46 percent of the time series examined. Random walk — undirected fluctuation without sustained directional trend — accounted for roughly 39 percent, and directional change (the pattern predicted by strict phyletic gradualism) was the best-fit model in only about 15 percent of cases.5 These quantitative findings broadly supported the punctuated equilibrium prediction that stasis is the predominant mode, while also confirming that evolutionary tempo is heterogeneous and that no single model describes all lineages.
Studies of individual clades have reinforced this picture. Satoshi Chiba's analysis of Mandarina land snails on the Bonin Islands documented both stasis within established species and rapid morphological shifts coinciding with colonization of new islands, consistent with the punctuated model in which isolation and novel environments trigger change.20 The bryozoan studies of Cheetham and Jackson demonstrated stasis over multimillion-year intervals with a statistical rigor that was difficult to attribute to preservational bias.14 Across diverse taxa — trilobites, bivalves, gastropods, foraminifera, bryozoans, mammals, and land snails — the accumulated evidence points to stasis as a genuine and pervasive feature of morphological evolution in the fossil record, not an artifact of incomplete sampling.5, 6
Species selection and macroevolution
If morphological change is concentrated at speciation events and species are stable between those events, then large-scale evolutionary trends cannot be explained solely by directional change within lineages. Instead, trends may result from the differential origination and extinction of species — a process analogous to natural selection but operating at the species level. Gould termed this mechanism species selection and argued that it was a logically necessary consequence of the punctuated equilibrium pattern.3, 15
For species selection to operate, species must possess heritable properties that influence their rates of speciation and extinction, just as organisms must possess heritable traits that influence their survival and reproduction for natural selection to operate. David Jablonski's extensive studies of Cretaceous and Cenozoic marine mollusks demonstrated that properties such as geographic range size and larval dispersal mode are indeed species-level traits (they characterize the species as a whole, not individual organisms) that influence extinction risk and speciation probability in predictable ways.15 Species with broad geographic ranges, for instance, tend to have lower extinction rates than geographically restricted species, and this differential survival can drive macroevolutionary trends even if no within-lineage directional change occurs.12, 15
The relationship between microevolution (changes within populations) and macroevolution (patterns above the species level) remains an active area of theoretical development. The strict hierarchical view advocated by Gould — that macroevolution is "decoupled" from microevolution and governed by its own distinct processes — has not been universally accepted. Many evolutionary biologists maintain that macroevolutionary patterns can in principle be fully explained by the accumulation of microevolutionary changes, even if the tempo is punctuated rather than gradual.7, 10 Nevertheless, the concept of species selection has been increasingly recognized as a legitimate and useful framework for understanding differential diversification, particularly in paleontological contexts where species-level data are more readily available than population-level data.15
Relationship to molecular evolution
Punctuated equilibrium was formulated as a theory about morphological evolution in the fossil record, but its relationship to molecular (DNA-sequence) evolution has been a recurrent question. Motoo Kimura's neutral theory of molecular evolution, proposed independently of punctuated equilibrium, holds that most molecular changes are selectively neutral and accumulate at a roughly constant rate (the molecular clock), driven by genetic drift rather than natural selection.16 If morphological stasis coexists with continuous molecular change, this would suggest that morphological evolution and molecular evolution are at least partially decoupled — a finding consistent with the idea that morphological stability is maintained by stabilizing selection or developmental constraints rather than by an absence of genetic variation.
Comparative studies have addressed whether molecular divergence between closely related species occurs gradually or in bursts associated with speciation. Some analyses of DNA sequence data have found elevated rates of molecular substitution along branches associated with speciation events, consistent with a punctuational model at the molecular level, while others have found no such acceleration.7 The question is complicated by the difficulty of distinguishing speciation-associated molecular change from the continuous background rate of neutral substitution, and by the different timescales over which morphological and molecular data are typically sampled. The current consensus is that molecular evolution is largely clock-like and not strongly punctuated, even in lineages whose morphological evolution is strikingly punctuated, reinforcing the view that morphological stasis reflects the action of stabilizing selection on phenotypes rather than an absence of underlying genetic change.7, 16
Current status and synthesis
More than five decades after its proposal, punctuated equilibrium occupies a settled if nuanced position within evolutionary biology. Its core empirical claims have been largely vindicated by quantitative paleontological research: stasis is real, pervasive, and not merely an artifact of gaps in the fossil record, and morphological change is often concentrated in association with lineage-splitting events.2, 5, 6 The theory's most important lasting contribution may be the recognition that stasis is a positive phenomenon requiring biological explanation, not a null expectation to be explained away.
At the same time, the debate has demonstrated that evolutionary tempo is heterogeneous. Neither strict punctuationism nor strict gradualism adequately describes the full range of evolutionary patterns observed across the tree of life. Some lineages show pronounced stasis interrupted by rapid change at speciation; others exhibit gradual directional trends; still others undergo random fluctuations without sustained direction or stability.5, 11 The relative frequency of these modes varies among taxa, traits, and environmental settings. Simpson's original insight — that evolution proceeds at variable rates and by variable modes — remains valid, enriched by the more precise quantitative framework that the punctuated equilibrium debate stimulated.7, 8
The hierarchical extensions of the theory, particularly species selection, continue to generate productive research in macroevolution and paleobiology. Jablonski's work on differential diversification in marine invertebrates has provided the strongest empirical support for species selection as a real macroevolutionary process.15 Meanwhile, the broader question of the relationship between micro- and macroevolution — whether large-scale patterns can be fully reduced to population-genetic processes or whether emergent properties at higher levels of the biological hierarchy play irreducible causal roles — remains one of the central unsettled questions in evolutionary theory.3, 12
Punctuated equilibrium did not overturn Darwinian evolution; it refined the discipline's understanding of how evolutionary change is distributed in time and across the biological hierarchy. By forcing biologists to confront the empirical reality of stasis and to take paleontological data seriously as evidence bearing on evolutionary process, Eldredge and Gould ensured that the fossil record would no longer be dismissed as too incomplete to inform evolutionary theory. In this respect, punctuated equilibrium succeeded in restoring paleontology to a central role in the study of evolution.2, 10
References
Phyletic size decrease in Hershey's kissing bugs? The case of Triatoma sanguisuga (Hemiptera: Reduviidae)
Microevolution and macroevolution: scale and hierarchy in evolutionary biology and paleobiology