Overview
- Anoxygenic photosynthesis arose in bacteria at least 3.5 billion years ago, using electron donors such as hydrogen sulphide and ferrous iron, while oxygenic photosynthesis — using water as the electron donor and releasing molecular oxygen — evolved in cyanobacteria and triggered the Great Oxidation Event approximately 2.4 billion years ago.
- Chloroplasts originated through the primary endosymbiotic capture of a cyanobacterium by a eukaryotic host cell approximately 1.5 billion years ago, and subsequent secondary and tertiary endosymbioses spread photosynthetic capacity to diverse eukaryotic lineages including diatoms, dinoflagellates, and brown algae.
- C4 photosynthesis, a carbon-concentrating mechanism that reduces photorespiration, evolved independently more than 60 times in flowering plants, making it one of the most striking examples of convergent evolution in the plant kingdom.
Photosynthesis — the conversion of light energy into chemical energy — is the biogeochemical process that sustains virtually all life on Earth, either directly through primary production or indirectly through the food webs that depend on it. The evolutionary history of photosynthesis spans more than three billion years and includes the origin of simple anoxygenic reaction centres in early bacteria, the invention of oxygenic photosynthesis by cyanobacteria that transformed Earth's atmosphere, the endosymbiotic incorporation of cyanobacteria into eukaryotic cells to form chloroplasts, and the repeated independent evolution of carbon-concentrating mechanisms in flowering plants.3, 4
{kind=link}
Origins of anoxygenic photosynthesis
The earliest photosynthetic organisms were bacteria that used light energy to drive electron transport chains but did not produce oxygen as a by-product. These anoxygenic phototrophs employed various inorganic and organic electron donors — including hydrogen sulphide, ferrous iron, and molecular hydrogen — rather than water, and they used either a Type I reaction centre (containing iron-sulphur clusters, as in green sulphur bacteria and heliobacteria) or a Type II reaction centre (containing a quinone acceptor, as in purple bacteria and green filamentous bacteria), but never both simultaneously.3, 11
Geological evidence suggests that photosynthesis was already operating by 3.5 billion years ago, and possibly earlier. Stromatolites — layered sedimentary structures formed by microbial mats — are found in rocks dating to approximately 3.5 billion years ago in the Pilbara Craton of Western Australia and the Barberton Greenstone Belt of South Africa, though whether the earliest stromatolites were produced by photosynthetic or chemotrophic organisms remains debated.16 Carbon isotope signatures in graphite from the Isua Supracrustal Belt of Greenland, dated to approximately 3.7 billion years ago, have been interpreted as evidence of biological carbon fixation, though these interpretations are also contested.5
The phylogenetic distribution of photosynthetic reaction centres across distantly related bacterial phyla — including Proteobacteria, Chlorobi, Chloroflexi, Firmicutes, and Acidobacteria — combined with the restricted occurrence of each reaction centre type to particular lineages, has led to the conclusion that the reaction centre genes have been distributed across bacterial phyla by horizontal gene transfer over billions of years.3, 11 The ancestral photosynthetic reaction centre was likely a homodimeric protein complex that predates the divergence of the Type I and Type II lineages.
The invention of oxygenic photosynthesis
Oxygenic photosynthesis, which uses water as the electron donor and releases molecular oxygen, evolved in the cyanobacterial lineage and represents one of the most consequential evolutionary innovations in Earth's history. The key innovation was the coupling of two photosystems in series: Photosystem II (PSII, a modified Type II reaction centre equipped with a manganese-containing oxygen-evolving complex capable of splitting water) and Photosystem I (PSI, a Type I reaction centre), linked by an electron transport chain. This tandem arrangement generates sufficient redox potential to extract electrons from water and use them to reduce NADP+, while simultaneously generating a proton gradient for ATP synthesis.3, 4
How the two photosystems came to be combined in a single organism remains one of the great questions in evolutionary biology. One hypothesis proposes that an ancestral organism possessing both Type I and Type II reaction centres (perhaps acquired by horizontal gene transfer) evolved the oxygen-evolving complex and linked the two systems, giving rise to the cyanobacterial lineage.11 Molecular clock estimates place the origin of oxygenic photosynthesis somewhere between 3.0 and 2.5 billion years ago, though some analyses suggest it may have evolved even earlier.1, 14
The consequences of oxygenic photosynthesis for Earth's biosphere were transformative. The accumulation of photosynthetically produced oxygen in the atmosphere led to the Great Oxidation Event (GOE) approximately 2.4 billion years ago, a transition recorded in multiple geochemical proxies including the disappearance of mass-independent fractionation of sulphur isotopes, the appearance of red beds (oxidised iron in terrestrial sediments), and the oxidation of detrital minerals such as pyrite and uraninite.2 The GOE fundamentally restructured global biogeochemical cycles, drove the evolution of aerobic metabolism, and enabled the subsequent evolution of complex multicellular life.2
Endosymbiotic origin of chloroplasts
The chloroplasts of eukaryotic algae and land plants originated through primary endosymbiosis: the engulfment and retention of a cyanobacterium by a heterotrophic eukaryotic host cell. Over evolutionary time, the endosymbiont lost most of its genes (transferred to the host nucleus) and became an obligate organelle, retaining only a small genome of approximately 100 to 200 genes that encodes core components of the photosynthetic machinery and the organellar gene expression apparatus.6
Multiple lines of evidence support the endosymbiotic origin of chloroplasts. Chloroplasts are bounded by a double membrane (consistent with engulfment by phagocytosis), possess their own circular DNA resembling cyanobacterial genomes, replicate by binary fission, and contain 70S ribosomes similar to those of bacteria rather than the 80S ribosomes of the eukaryotic cytoplasm.6 Phylogenomic analyses have consistently placed chloroplast genomes within the cyanobacterial radiation, confirming their cyanobacterial ancestry.8
Current evidence strongly supports a single primary endosymbiotic event giving rise to all primary plastids, which are found in three eukaryotic lineages: the Glaucophyta, Rhodophyta (red algae), and Viridiplantae (green algae and land plants). These three groups together form the supergroup Archaeplastida.8 The glaucophyte plastid (called a cyanelle) retains a vestigial peptidoglycan cell wall between its two membranes, a remarkable living intermediate that preserves a relic of the original cyanobacterial endosymbiont's bacterial cell wall.6
Secondary and tertiary endosymbioses
Photosynthetic capacity spread far beyond the Archaeplastida through secondary endosymbiosis, in which a heterotrophic eukaryote engulfed a photosynthetic eukaryote (a red or green alga) and retained its plastid. Secondary endosymbiotic plastids are characterised by additional membranes (three or four, reflecting the successive engulfment events) and, in some cases, a remnant of the algal nucleus called a nucleomorph.12
Secondary endosymbiosis involving red algae gave rise to the plastids of heterokonts (diatoms, brown algae, chrysophytes), haptophytes, cryptophytes, and dinoflagellates — collectively responsible for a major fraction of marine primary productivity.12 Secondary endosymbiosis involving green algae gave rise to the plastids of euglenids and chlorarachniophytes. In some dinoflagellate lineages, further rounds of endosymbiosis (tertiary endosymbiosis) have occurred, in which the original red algal-derived plastid was replaced by a plastid obtained from a haptophyte, diatom, or other photosynthetic eukaryote. These serial endosymbioses demonstrate that the acquisition of photosynthetic capacity is a recurrent theme in eukaryotic evolution, and that massive horizontal gene transfer from endosymbiont to host has been a recurring feature of eukaryotic genome evolution.7, 12, 17
RuBisCO and the C3 limitation
The enzyme ribulose-1,5-bisphosphate carboxylase/oxygenase (RuBisCO) catalyses the first step of carbon fixation in the Calvin-Benson cycle, the primary pathway of CO2 assimilation in photosynthetic organisms. RuBisCO is one of the most abundant proteins on Earth, but it is also one of the slowest and least specific: it has a catalytic rate of only a few reactions per second and cannot perfectly distinguish between CO2 and O2 as substrates.15 When RuBisCO fixes O2 instead of CO2, it initiates the photorespiratory pathway, which consumes energy and releases previously fixed carbon, reducing the net efficiency of photosynthesis by 20 to 50 percent under warm, dry conditions.15
The oxygenase activity of RuBisCO is an evolutionary legacy of its origin in an anoxic world. When RuBisCO first evolved, atmospheric O2 levels were negligible and the enzyme's inability to discriminate between CO2 and O2 was irrelevant. As oxygenic photosynthesis raised atmospheric O2 concentrations, the oxygenase activity became an increasing liability, particularly under conditions of high temperature and low CO2 that favour the oxygenase reaction.15 This limitation has imposed strong selective pressure on photosynthetic organisms to evolve mechanisms that concentrate CO2 around RuBisCO, suppressing its oxygenase activity.
The evolution of C4 photosynthesis
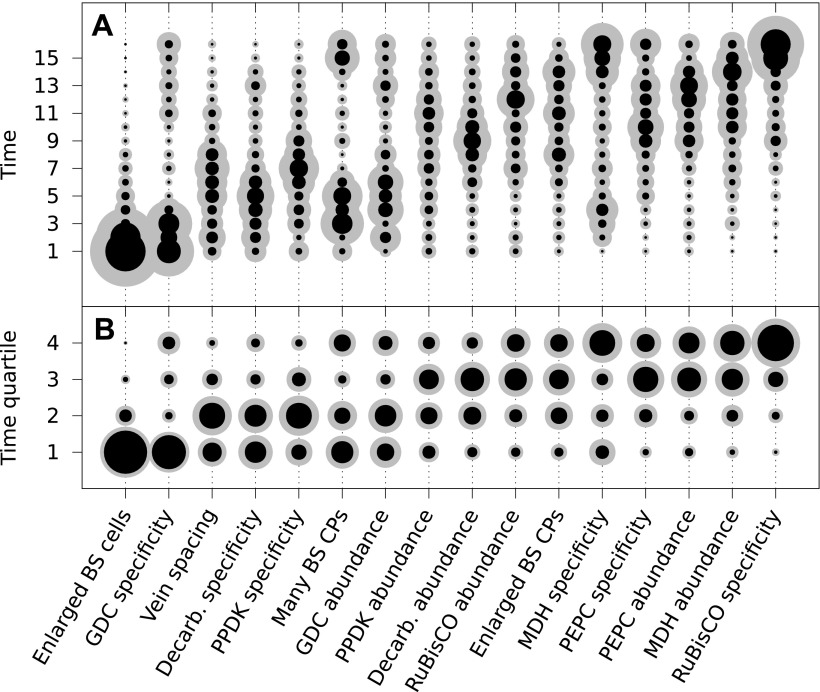
C4 photosynthesis is a carbon-concentrating mechanism in which CO2 is initially fixed by the enzyme phosphoenolpyruvate carboxylase (PEPC) in mesophyll cells to form a four-carbon acid (hence "C4"), which is then transported to bundle sheath cells where it is decarboxylated, releasing CO2 at high concentration around RuBisCO. This spatial separation of initial carbon fixation and the Calvin cycle effectively eliminates photorespiration and confers a significant advantage in hot, dry, high-light environments.9, 10
C4 photosynthesis has evolved independently at least 66 times across 19 families of flowering plants, making it one of the most remarkable examples of convergent evolution in biology.10 Most C4 origins occurred within the grasses (Poaceae) and the sedges (Cyperaceae), but independent origins are also found in eudicot families including Chenopodiaceae, Euphorbiaceae, and Asteraceae. The repeated convergent evolution of C4 photosynthesis suggests that the genetic and anatomical prerequisites for this pathway — including Kranz anatomy (a wreath-like arrangement of bundle sheath cells around veins) and the regulatory elements controlling PEPC expression — were already present in C3 ancestors, requiring relatively few evolutionary steps to assemble into a functional C4 pathway.9, 10
The ecological expansion of C4 grasslands approximately 6 to 8 million years ago, during the late Miocene, was a major event in terrestrial ecosystem history. Stable carbon isotope records from paleosols, fossil teeth, and marine sediments document a global shift from C3-dominated to C4-dominated grassland ecosystems during this period, likely driven by declining atmospheric CO2 levels, increased fire frequency, and seasonal aridity.10
Crassulacean acid metabolism
Crassulacean acid metabolism (CAM) is a second major carbon-concentrating mechanism that evolved independently of C4 photosynthesis. In CAM plants, CO2 is fixed by PEPC at night, when stomata are open and transpirational water loss is minimised, and stored as malic acid in vacuoles. During the day, the stomata close, the stored malic acid is decarboxylated, and the released CO2 is refixed by RuBisCO in the Calvin cycle. CAM thus achieves temporal rather than spatial separation of initial CO2 fixation and the Calvin cycle.13
CAM has evolved independently in over 35 families of vascular plants, including cacti, agaves, bromeliads, orchids, and stonecrops, and is found in approximately 6 percent of flowering plant species.13, 18 Like C4 photosynthesis, the repeated convergent evolution of CAM implies that the underlying enzymatic and regulatory components were available in ancestral C3 plants and required relatively modest modifications to be assembled into a functional CAM pathway. Both CAM and C4 photosynthesis illustrate how natural selection can repeatedly arrive at similar biochemical solutions when organisms face analogous environmental challenges.10, 13
References
Primary endosymbiosis and the evolution of light and oxygen sensing in photosynthetic eukaryotes
Acquisition of 1,000 eubacterial genes physiologically transformed a methanogen at the origin of Haloarchaea