Overview
- Phenotypic plasticity is the ability of a single genotype to produce different phenotypes in response to different environmental conditions, described mathematically by reaction norms that map the range of phenotypic expression across an environmental gradient.
- Classic examples include predator-induced helmet formation in Daphnia, temperature-dependent sex determination in reptiles, and seasonal polyphenism in butterflies — all demonstrating that organisms are not passive expressions of their genes but active responders to environmental cues.
- The plasticity-first evolution hypothesis proposes that environmentally induced phenotypic changes can precede and facilitate genetic adaptation by exposing novel phenotypes to selection, with genetic assimilation subsequently stabilising the initially plastic trait.
Phenotypic plasticity is the ability of a single genotype to produce different phenotypes in response to different environmental conditions. Rather than being a fixed blueprint that specifies a single invariant outcome, a genotype encodes a range of potential phenotypic expressions, and the environment experienced during development determines which phenotype is realised. The concept was first articulated by Richard Woltereck in 1909, who coined the term Reaktionsnorm (reaction norm) to describe the function mapping a genotype's phenotypic expression across an environmental gradient while studying head morphology in Daphnia.1 Phenotypic plasticity encompasses phenomena as diverse as predator-induced morphological defences, temperature-dependent sex determination, seasonal colour changes, and behavioural flexibility, and it plays a fundamental role in how organisms cope with environmental heterogeneity, colonise novel habitats, and potentially initiate new evolutionary trajectories.2, 10
{kind=link}
Reaction norms and the scope of plasticity
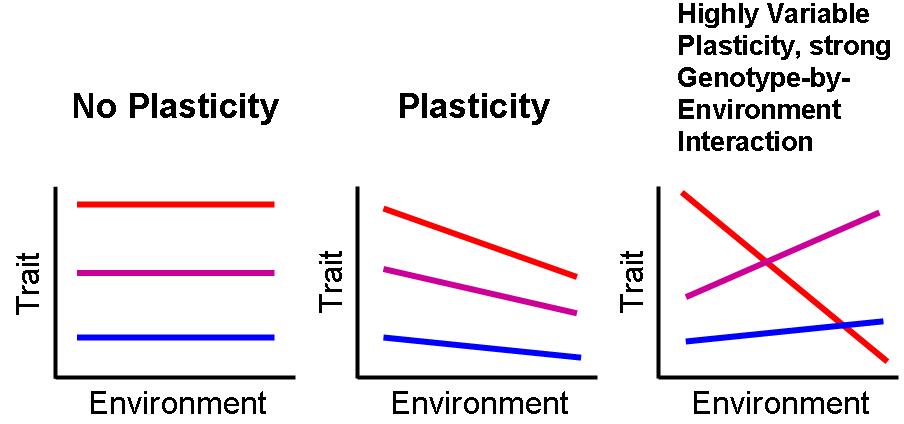
The reaction norm is the foundational concept for understanding phenotypic plasticity. It describes the set of phenotypes that a particular genotype produces across a range of environments, and it can be visualised as a curve on a graph where the x-axis represents the environmental variable and the y-axis represents the phenotypic value. A flat reaction norm indicates a genotype that produces the same phenotype regardless of environment (canalization), while a steep reaction norm indicates high plasticity. The shape of the reaction norm, including whether it is linear or nonlinear, monotonic or U-shaped, may itself vary among genotypes, a phenomenon termed genotype-by-environment interaction (G × E).2, 5
Plasticity manifests at multiple levels of biological organisation. At the molecular level, gene expression patterns change in response to environmental signals such as temperature, photoperiod, nutrient availability, and the presence of predators or pathogens. At the cellular level, these expression changes can alter cell differentiation, proliferation, and apoptosis. At the organismal level, the consequences range from subtle quantitative shifts in body size or metabolic rate to dramatic qualitative transformations in morphology, life history, or behaviour. The most extreme form of discrete plasticity is polyphenism, in which a single genotype produces two or more distinct phenotypic forms (morphs) depending on environmental cues, as seen in the caste systems of social insects or the seasonal wing patterns of butterflies.3, 10
Adaptive versus non-adaptive plasticity
Not all plasticity is adaptive. Adaptive plasticity occurs when the environmentally induced phenotypic change improves the organism's fitness in the environment that triggered it: a tadpole that develops a deeper tail fin in the presence of dragonfly predators swims faster and survives better than one that does not.11 Non-adaptive plasticity, by contrast, is a passive or pathological response to environmental stress that does not improve fitness, such as stunted growth caused by malnutrition or developmental abnormalities caused by extreme temperatures. The distinction is important because adaptive plasticity implies that the plastic response has been shaped by natural selection, with the reaction norm itself being the target of selection, whereas non-adaptive plasticity is simply a consequence of the physical and biochemical constraints of development.2, 4
Ghalambor and colleagues provided empirical evidence that both adaptive and non-adaptive plasticity can play roles in evolution. In their 2015 study of Trinidadian guppies transplanted to novel environments, they found that gene expression changes in the new environment were a mixture of adaptive plasticity (expression changes in the direction favoured by natural selection) and non-adaptive plasticity (expression changes in the opposite direction). Notably, the non-adaptive plastic responses showed the strongest signatures of subsequent genetic evolution, suggesting that maladaptive plasticity can create strong selection pressure that drives rapid evolutionary change.19
Classic examples
The water flea Daphnia has served as a model system for the study of phenotypic plasticity since Woltereck's original work over a century ago. When exposed to chemical cues (kairomones) released by predators such as the phantom midge larva Chaoborus, Daphnia species develop defensive morphological structures including elongated head helmets, neck teeth, and tail spines that reduce their vulnerability to predation. These structures are not produced in the absence of predator cues, demonstrating that the response is an environmentally triggered developmental programme rather than a constitutive trait. Tollrian and Harvell documented the molecular and ecological mechanisms underlying these inducible defences, showing that the plastic response is mediated by specific chemoreceptors that detect predator-derived molecules and activate downstream developmental pathways.12
Tadpole morphology provides another extensively studied example. McCollum and Van Buskirk demonstrated that tadpoles of several frog species develop dramatically different body shapes depending on whether they are reared in the presence or absence of predators. In predator environments, tadpoles develop deeper tail fins and smaller bodies, which increase swimming speed and escape ability at the cost of reduced competitive feeding efficiency. In predator-free environments, the same genotypes produce shallow-tailed, large-bodied tadpoles that are better competitors for food but more vulnerable to predation. The plasticity is induced by chemical cues from predators and is reversible to some degree if the cues are removed, demonstrating remarkable developmental flexibility.11
Temperature-dependent sex determination (TSD) in reptiles is a particularly striking form of plasticity in which the incubation temperature of eggs determines the sex of the developing embryo. In many turtle species, low temperatures produce males and high temperatures produce females, while in some crocodilians and lizards the pattern is reversed or more complex. Janzen and Phillips reviewed the evolution and ecological significance of TSD and noted that the phenomenon is widespread across reptilian lineages, suggesting either deep homology or repeated evolution. The fitness consequences of TSD are debated, but one leading hypothesis proposes that it is adaptive when the incubation environment differentially affects male and female fitness, so that each sex is produced under the conditions most favourable for its reproductive success.13
Seasonal polyphenism in butterflies demonstrates how environmental cues experienced during development can produce dramatically different adult phenotypes. In the African butterfly Bicyclus anynana, individuals that develop during the wet season produce wing patterns with prominent eyespots that deflect predator attacks toward the wing margins, while dry-season individuals produce cryptic wing patterns that provide camouflage against dead leaf litter. Brakefield and colleagues showed that the switch between morphs is triggered by the temperature experienced during a critical period of larval development, with specific hormonal pathways mediating the developmental response.17
Genetic accommodation and assimilation
The relationship between phenotypic plasticity and genetic evolution was explored experimentally by Conrad Hal Waddington in a series of landmark experiments in the 1940s and 1950s. Waddington exposed Drosophila melanogaster pupae to heat shock and observed that a small proportion developed a crossveinless wing phenotype. By selecting for this phenotype each generation and continuing the heat-shock treatment, he found that the proportion of crossveinless individuals increased over generations. Crucially, after several generations of selection, some individuals produced the crossveinless phenotype even without heat shock, demonstrating that an initially environmentally induced trait had become genetically fixed. Waddington termed this process genetic assimilation.6, 7
Genetic assimilation is now understood as one outcome of the broader process of genetic accommodation, a term introduced by Mary Jane West-Eberhard. Genetic accommodation occurs when a novel phenotype, initially induced by an environmental change, is subsequently refined by natural selection acting on heritable variation in the form and regulation of the plastic response. The result can be either genetic assimilation, in which the trait becomes constitutively expressed and loses its environmental dependence, or genetic adaptation of the plasticity itself, in which the reaction norm is refined to produce a more finely tuned response to the environmental cue.9, 10
Ledon-Rettig and colleagues provided empirical evidence for genetic accommodation in the wild by studying spadefoot toad tadpoles (Spea). Tadpoles of some species develop either an omnivore morph or a carnivore morph depending on the availability of shrimp prey. The carnivore morph, characterised by a wider head and enlarged jaw muscles, is initially induced by dietary cues but shows evidence of genetic accommodation: in populations that have experienced long-term exposure to the alternative diet, the threshold for inducing the carnivore morph has shifted, and the morphological differences between morphs have become more pronounced, consistent with selection refining the plastic response over evolutionary time.8
Plasticity-first evolution
The plasticity-first evolution hypothesis, articulated by Levis and Pfennig, extends the concept of genetic accommodation into a broader framework for understanding the role of plasticity in generating evolutionary novelty. The hypothesis proposes that when organisms encounter a novel environment, phenotypic plasticity provides the initial means of coping, producing new phenotypes that expose the population to new selective pressures. If the plastic response is at least partially adaptive, it allows the population to persist in the new environment, buying time for natural selection to act on heritable variation in the reaction norm and to genetically stabilise or refine the initially plastic trait. In this way, plasticity does not merely allow organisms to tolerate environmental change but actively initiates new evolutionary trajectories by revealing phenotypic variation that selection can act upon.15, 16
Price, Qvarnström, and Irwin provided theoretical support for this view, arguing that phenotypic plasticity can drive genetic evolution through several mechanisms: it can expose hidden genetic variation to selection by altering the phenotypic expression of existing alleles, it can shift populations to new positions in the fitness landscape where different selective pressures operate, and it can facilitate the colonisation of new environments that then impose novel selection regimes. Their model predicts that plasticity should be most important in driving evolution during the early stages of adaptation to a new environment, when the population has not yet had time to evolve genetically optimised responses.20
Costs and limits of plasticity
If plasticity allows organisms to produce the optimal phenotype for every environment, one might expect all organisms to be infinitely plastic. The fact that they are not implies that plasticity carries costs and is subject to limits. DeWitt, Sih, and Wilson identified several categories of costs associated with phenotypic plasticity. Maintenance costs arise from the sensory, neural, and regulatory machinery required to detect environmental cues and implement developmental responses. Production costs occur when producing a phenotype through a plastic pathway is more expensive than producing the same phenotype through a constitutive pathway. Information reliability costs arise when environmental cues are unreliable predictors of future conditions, causing the organism to produce a phenotype mismatched to its actual selective environment. Developmental instability costs may arise if the regulatory flexibility required for plasticity reduces the precision of developmental processes.4
Limits on plasticity are distinct from costs. A limit is a constraint on the range or accuracy of plastic responses, even if the organism has been selected for plasticity. Developmental time lags mean that plastic responses cannot be instantaneous; an organism that takes weeks to develop an induced defence may be killed by a predator in the interim. Irreversibility constrains organisms whose developmental programmes cannot be reversed once initiated, as in temperature-dependent sex determination, where the sex of a reptile is fixed once the gonads have differentiated, regardless of subsequent temperature changes. Spatial heterogeneity at scales finer than the organism's ability to detect or respond to creates additional limits, as does the reliability of environmental cues as predictors of future conditions.2, 4
Role in colonisation of novel environments
Phenotypic plasticity has been proposed as a key trait facilitating the colonisation of novel environments, because plastic organisms can adjust their phenotype to match local conditions without requiring immediate genetic adaptation. Agrawal argued that species with greater phenotypic plasticity should be more successful colonisers because they can maintain fitness across a broader range of environmental conditions, and empirical studies of invasive species have generally supported this prediction.18
Hendry, Farrugia, and Kinnison reviewed the role of plasticity in responses to human-caused environmental change and found that many populations initially respond to novel anthropogenic stressors through plasticity rather than genetic adaptation. Urban heat islands, altered photoperiods from artificial light, novel food sources, and chemical pollutants all induce plastic responses in exposed populations. Whether these plastic responses are ultimately adaptive, allowing populations to persist until genetic adaptation can catch up, or maladaptive, accelerating population decline, depends on the specific organisms and stressors involved. The distinction has important implications for conservation biology, because managing for plasticity (maintaining the conditions that allow plastic responses to be expressed) may be as important as managing for genetic diversity in populations facing rapid environmental change.14
Significance
Phenotypic plasticity challenges the view of organisms as passive products of their genotypes and environments, revealing instead that development is an active process in which environmental information is integrated with genetic programmes to produce context-dependent phenotypes. The recognition that plasticity is itself a heritable, evolvable trait subject to natural selection has broadened the understanding of how organisms adapt to heterogeneous and changing environments, and the plasticity-first evolution framework has reinvigorated debates about the relationship between development and evolution that date back to the pre-Modern Synthesis era.9, 10, 16
The practical relevance of plasticity extends from agriculture, where crop breeders must account for genotype-by-environment interactions in selecting varieties for diverse growing conditions, to medicine, where developmental plasticity underlies the "thresholds" and critical periods that shape disease susceptibility, to conservation biology, where the capacity for plastic responses may determine which species can tolerate rapid climate change and which cannot. Understanding the mechanisms, evolution, and limits of phenotypic plasticity remains one of the central challenges of modern evolutionary and developmental biology.2, 3, 14
References
Weitere experimentelle Untersuchungen über Artänderung, speziel über das Wesen quantitativer Artunterschiede bei Daphniden
Ecological developmental biology: the environmental regulation of development, health, and evolution
The ecology and genetics of fitness in Chlamydomonas. X. The analysis of genotype-environment interaction
Genetic accommodation in the wild: evolution of gene expression plasticity during character displacement
Predator-induced morphological changes in an amphibian: predation by dragonflies affects tadpole shape and color
Plasticity-first evolution: the role of phenotypic plasticity in the evolution of novelty
Seasonal polyphenism in butterflies: a case study of the African butterfly Bicyclus anynana