Overview
- Viruses evolve at rates orders of magnitude faster than cellular organisms, with RNA viruses mutating at approximately 10⁻³ to 10⁻⁵ substitutions per nucleotide per replication cycle — a million-fold higher than most DNA-based genomes — enabling them to adapt to host defences, antiviral drugs, and new host species on timescales of weeks to months.
- Manfred Eigen's quasispecies theory describes viral populations not as collections of identical genomes but as dynamic mutant clouds clustered around a master sequence, where the population's collective genetic diversity rather than any single genome determines evolutionary trajectory and where exceeding an error threshold causes irreversible loss of genetic information.
- Host-switching events, in which a virus adapts to infect a new host species, have produced some of the most consequential disease emergence events in human history, including the HIV pandemic (from chimpanzee SIV), the 1918 influenza pandemic (from avian reservoirs), and the SARS-CoV-2 pandemic (likely from a bat coronavirus ancestor).
Viruses are the most abundant biological entities on Earth and among the fastest-evolving, with mutation rates, generation times, and population sizes that compress into months the evolutionary dynamics that take millennia in cellular organisms. Their rapid evolution underlies some of the most pressing challenges in medicine and public health, from the annual recurrence of influenza epidemics to the emergence of drug-resistant HIV and the global spread of novel coronaviruses.3, 4 The study of viral evolution draws on population genetics, molecular biology, and epidemiology to understand how viral populations generate and maintain genetic diversity, how they adapt to host immune responses and antiviral drugs, and how they occasionally cross species barriers to cause pandemics.15
{kind=link}
Mutation rates and error-prone replication
The defining feature of viral evolution is the extraordinary mutation rate of viral genomes. RNA viruses, which include influenza, HIV, Ebola, and the coronaviruses, typically mutate at rates of approximately 10−3 to 10−5 substitutions per nucleotide site per round of genome replication, roughly a million times faster than most DNA-based cellular organisms.4 These high rates arise because RNA-dependent RNA polymerases and reverse transcriptases lack the proofreading exonuclease activity that corrects errors during DNA replication in cellular organisms. The result is that every replication cycle produces a population of genomes containing numerous mutations relative to the parental template.3, 5
DNA viruses generally have lower mutation rates than RNA viruses, typically in the range of 10−6 to 10−8 substitutions per site per replication, because many DNA viruses encode or co-opt host proofreading polymerases.4 An important exception among RNA viruses is the coronaviruses (family Coronaviridae), which possess a 3′-to-5′ exoribonuclease encoded by the nsp14 gene that provides a rudimentary proofreading function. This proofreading lowers their mutation rate to approximately 10−6 substitutions per site per replication cycle, among the lowest of any RNA virus, and may be one reason coronaviruses can maintain genomes of approximately 30 kilobases — the largest of any RNA virus — without suffering error catastrophe.5, 12
Quasispecies theory
The quasispecies concept, developed by Manfred Eigen in 1971 and later elaborated by Eigen and Peter Schuster, provides a theoretical framework for understanding the population structure of rapidly mutating replicators. Rather than viewing a viral population as a collection of identical genomes or even a set of discrete variants, quasispecies theory describes the population as a dynamic "mutant cloud" of closely related but non-identical sequences clustered in sequence space around a central master sequence.1, 2 The evolutionary unit in this framework is not the individual genome but the entire distribution of mutant sequences, which collectively determines the population's phenotypic properties and evolutionary trajectory.2, 3
A key prediction of quasispecies theory is the existence of an error threshold: a maximum mutation rate above which the genetic information encoded in the master sequence can no longer be maintained by selection. If the mutation rate exceeds this threshold, the population loses its coherence around the master sequence and drifts randomly through sequence space, effectively losing its genetic identity. This prediction has motivated the development of lethal mutagenesis as an antiviral strategy, in which mutagenic drugs push viral populations past their error threshold, causing an irreversible loss of fitness and eventual population collapse. Proof-of-concept experiments have demonstrated that nucleoside analogues such as ribavirin and favipiravir can indeed increase the mutation rate of RNA viruses to lethal levels in vitro.6
Recombination and reassortment
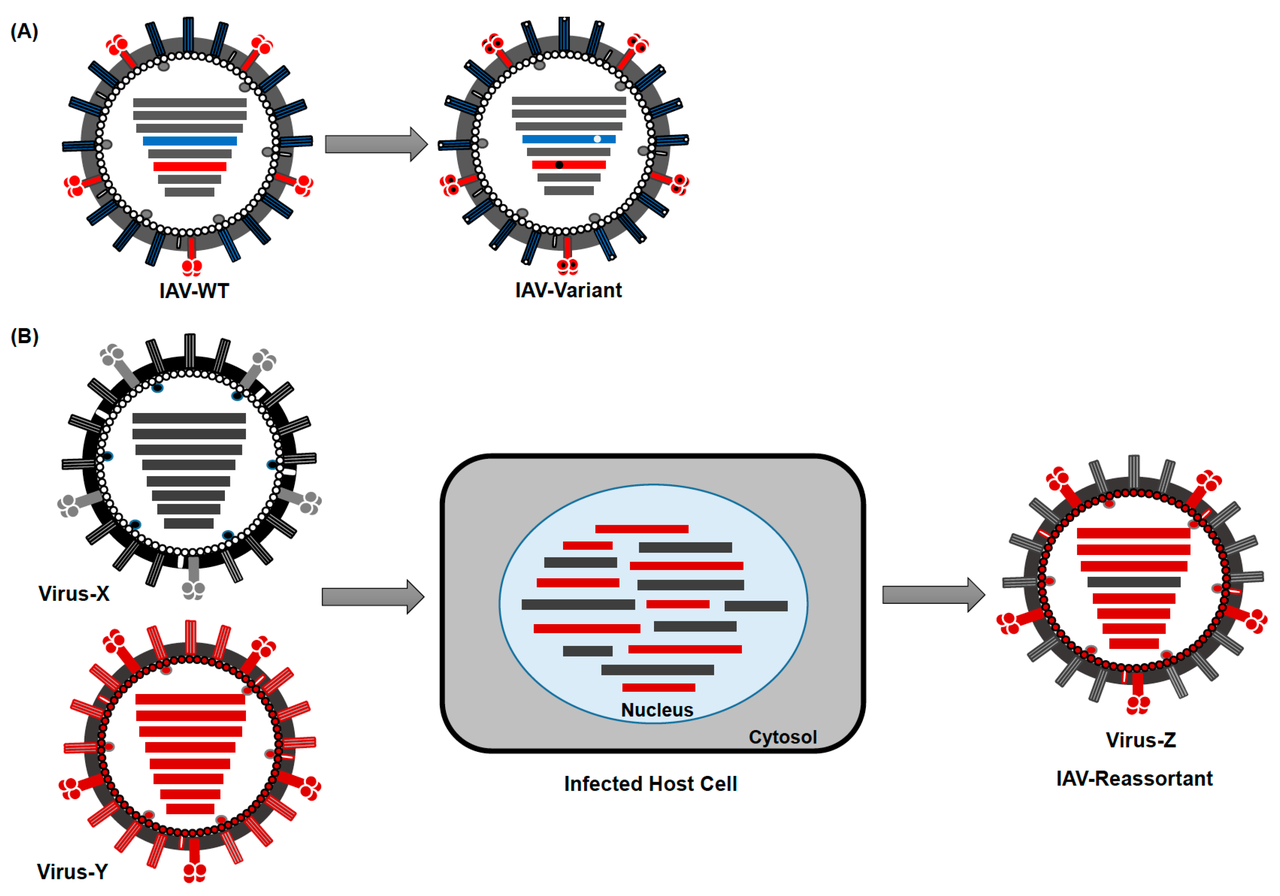
In addition to mutation, viral genomes generate diversity through recombination (exchange of genetic material between co-infecting viral genomes) and, in segmented viruses, reassortment (exchange of whole genome segments between co-infecting viruses). Recombination occurs in many RNA virus families and is especially frequent in coronaviruses, where the polymerase frequently jumps between templates during replication, a process known as copy-choice recombination. This mechanism has been implicated in the generation of novel coronaviruses with altered host range, including the likely origin of SARS-CoV-2's receptor-binding domain through ancestral recombination events among bat sarbecoviruses.12, 13
Reassortment is the primary mechanism of genetic exchange in influenza viruses, whose genomes consist of eight separate RNA segments. When two different influenza strains co-infect the same cell, progeny virions can package segments from both parents, generating novel combinations. This process, termed antigenic shift, can produce pandemic strains against which the human population has no pre-existing immunity. The 1957 Asian influenza and 1968 Hong Kong influenza pandemics both arose from reassortment events between human and avian influenza viruses, combining avian haemagglutinin genes with the internal gene segments of circulating human strains.14, 16
Host-switching and pandemic emergence
Host-switching events, in which a virus adapts to infect and transmit within a new host species, have produced some of the most consequential epidemics and pandemics in human history. The adaptation of a chimpanzee simian immunodeficiency virus (SIVcpz) to humans gave rise to HIV-1, the causative agent of the AIDS pandemic. Phylogenetic analyses by Gao and colleagues, and subsequently by Keele and colleagues, established that HIV-1 group M — responsible for the vast majority of global infections — originated from a single cross-species transmission event from the chimpanzee subspecies Pan troglodytes troglodytes in west-central Africa, likely in the early twentieth century.7, 8
Influenza provides a recurring example of host-switching. The 1918 pandemic, caused by an H1N1 virus, killed an estimated 50 million people worldwide and arose from an avian-origin virus that adapted to efficient human-to-human transmission.9 Reconstruction of the 1918 virus by Tumpey and colleagues confirmed its exceptional virulence and transmissibility in animal models, and subsequent phylogenetic work has traced the evolutionary relationships among avian, swine, and human influenza lineages, revealing a complex network of cross-species transmission and adaptation.9, 14
The emergence of SARS-CoV-2 in late 2019 demonstrated the pandemic potential of coronaviruses. Genomic analysis by Zhou and colleagues showed that SARS-CoV-2 shares 96.2% genome-wide nucleotide identity with bat coronavirus RaTG13, indicating a likely origin from bat sarbecovirus reservoirs, though the precise pathway of transmission to humans — whether directly from bats or through an intermediate animal host — remains under investigation.11, 12 The subsequent evolution of SARS-CoV-2 within the human population has been extensively documented, including the rapid global spread of the D614G spike mutation, which increased viral infectivity, and the successive emergence of variants of concern (Alpha, Delta, Omicron) bearing mutations in the spike protein that enhanced transmissibility or partially evaded vaccine-induced immunity.18
Drug resistance and immune evasion
The rapid evolution of viruses poses a direct challenge to both antiviral therapy and vaccination. HIV-1 provides the paradigmatic case of drug-resistance evolution: the virus's high mutation rate, enormous within-host population size (approximately 1010 virions produced per day in an untreated patient), and short generation time mean that resistance mutations to any single antiretroviral drug emerge within weeks of monotherapy. This observation drove the development of highly active antiretroviral therapy (HAART), which combines three or more drugs targeting different viral proteins, making it statistically improbable that a single virus would simultaneously acquire resistance mutations to all drugs in the regimen.17
Influenza virus evades population-level immunity through antigenic drift, the gradual accumulation of amino acid substitutions in the surface glycoproteins haemagglutinin (HA) and neuraminidase (NA) that alter the antigenic properties of the virus. Smith and colleagues used antigenic cartography to map the antigenic evolution of influenza A H3N2 over 35 years, revealing that antigenic change occurs in a punctuated pattern: periods of relative antigenic stasis are interrupted by abrupt shifts to new antigenic clusters, each of which triggers a new epidemic wave.10 This pattern of antigenic evolution necessitates the annual reformulation of influenza vaccines and has motivated the search for broadly neutralising antibodies and "universal" vaccines targeting conserved epitopes.10, 14
The study of viral evolution illuminates evolutionary processes at an accelerated pace and with a resolution rarely achievable in other systems. The molecular phylodynamics framework, which integrates phylogenetic, epidemiological, and population-genetic analyses, has enabled researchers to reconstruct the transmission histories of outbreaks in near-real time, to quantify the rate at which natural selection drives antigenic change, and to predict the future trajectory of viral evolution with increasing accuracy.15 As new viruses continue to emerge from animal reservoirs and established viruses continue to evolve resistance to drugs and vaccines, understanding viral evolution remains one of the most consequential applications of evolutionary biology to human welfare.
References
Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic
Recombination, reservoirs, and the modular spike: mechanisms of coronavirus cross-species transmission