Overview
- Richard Wrangham's cooking hypothesis argues that the control of fire and habitual cooking were not merely useful technologies but the single most transformative dietary transition in hominin evolution, directly driving the anatomical and neurological changes that distinguish Homo erectus from its australopith predecessors roughly 1.9 million years ago.
- Experimental evidence demonstrates that cooking increases the net caloric yield of starch-rich foods by 30–100% through gelatinization and increases the digestible energy from meat through protein denaturation, providing the energetic surplus that could have sustained a dramatically expanded brain and reduced gut.
- The hypothesis faces a significant evidentiary gap: secure archaeological evidence for controlled fire use extends back only to roughly 1 million years ago at Wonderwerk Cave, South Africa, and habitual fire use is firmly established only from about 400,000–350,000 years ago, creating a nearly million-year gap between the predicted onset of cooking and the earliest uncontested fire evidence.
The cooking hypothesis is the proposal that the habitual use of fire to cook food was a transformative event in hominin evolution, one that drove major anatomical, neurological, and social changes in the lineage leading to modern humans. Most closely associated with the primatologist and biological anthropologist Richard Wrangham of Harvard University, the hypothesis argues that cooking was not merely a useful technological innovation but the single most consequential dietary transition in the human past, directly responsible for the suite of changes that distinguish Homo erectus from its australopith predecessors.1, 2 The hypothesis integrates evidence from nutritional science, comparative anatomy, primate ecology, and archaeology to argue that cooking fundamentally reshaped the human body, brain, and social life. It remains one of the most debated and productive frameworks in the study of human evolution.
Wrangham's thesis
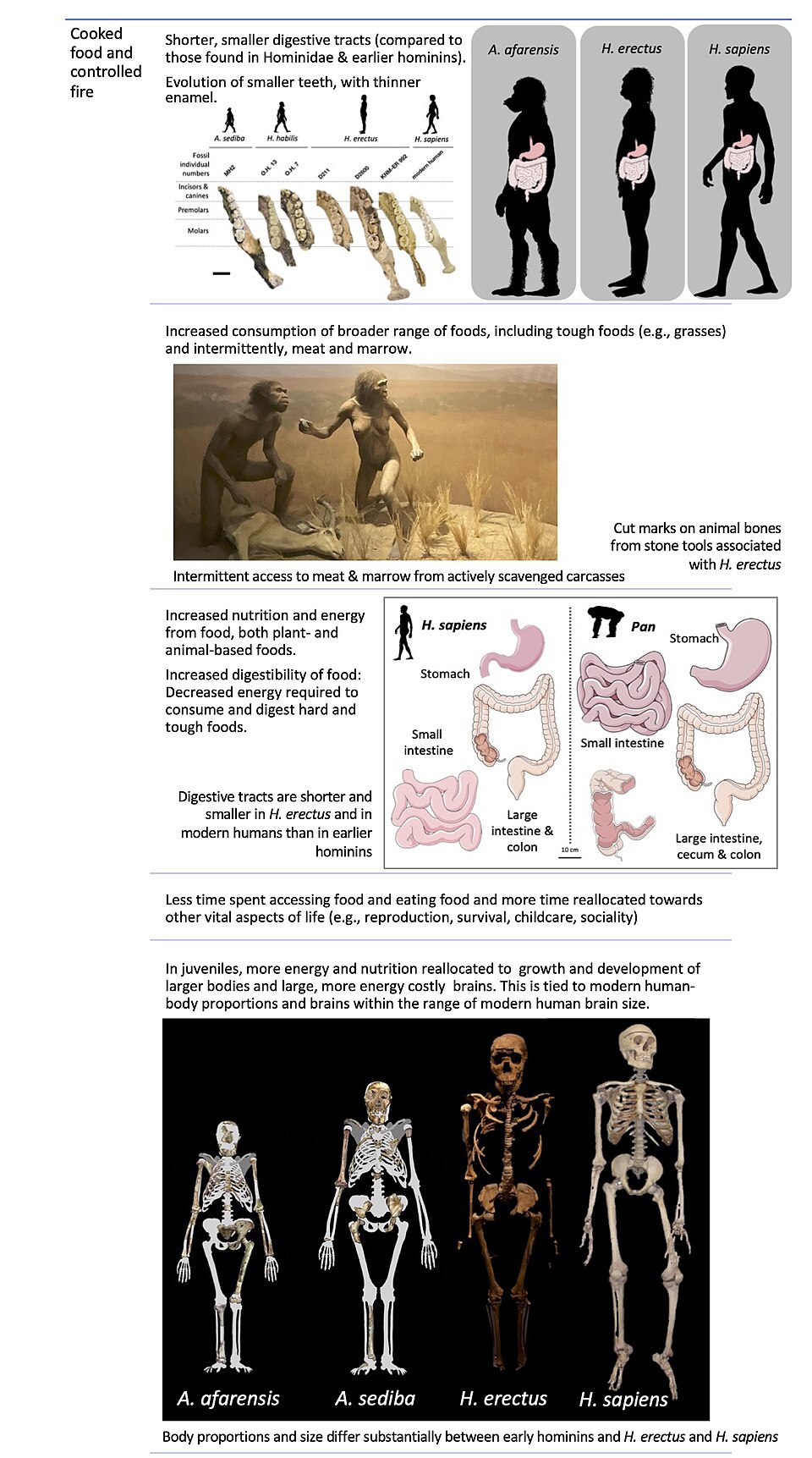
The cooking hypothesis was first articulated in a 1999 paper by Wrangham and colleagues in Current Anthropology, titled "The Raw and the Stolen," and was elaborated at length in Wrangham's 2009 book Catching Fire: How Cooking Made Us Human.1, 2 The argument begins from a simple observation: cooking dramatically alters the physical and chemical properties of food in ways that increase its caloric yield and reduce the mechanical and metabolic costs of digestion. Wrangham contended that these effects are not marginal supplements to an otherwise adequate raw diet but rather represent a qualitative shift in the energetic returns available to a hominin that mastered fire. In his formulation, cooking provided the energetic surplus that made possible the enlargement of the brain, the reduction of the gut, the gracilization of the teeth and jaws, and the reorganization of social life around the campfire.1
The core logic of the hypothesis rests on a well-established metabolic constraint. The human brain consumes approximately 20–25% of the body's resting energy despite constituting only about 2% of body mass, a metabolic load far greater than that of any other primate.16 In 1995, Leslie Aiello and Peter Wheeler proposed the expensive tissue hypothesis, arguing that the enlarged human brain was financed not by an increase in total metabolic rate but by a compensatory reduction in the gastrointestinal tract, another metabolically expensive organ system.16 Wrangham seized on this framework and proposed a specific mechanism: cooking was the dietary innovation that allowed the gut to shrink. By softening food and gelatinizing starches, cooking externalized much of the mechanical and chemical work of digestion, reducing the need for a large, energy-intensive gut while simultaneously increasing the net energy extracted from each meal.1, 5
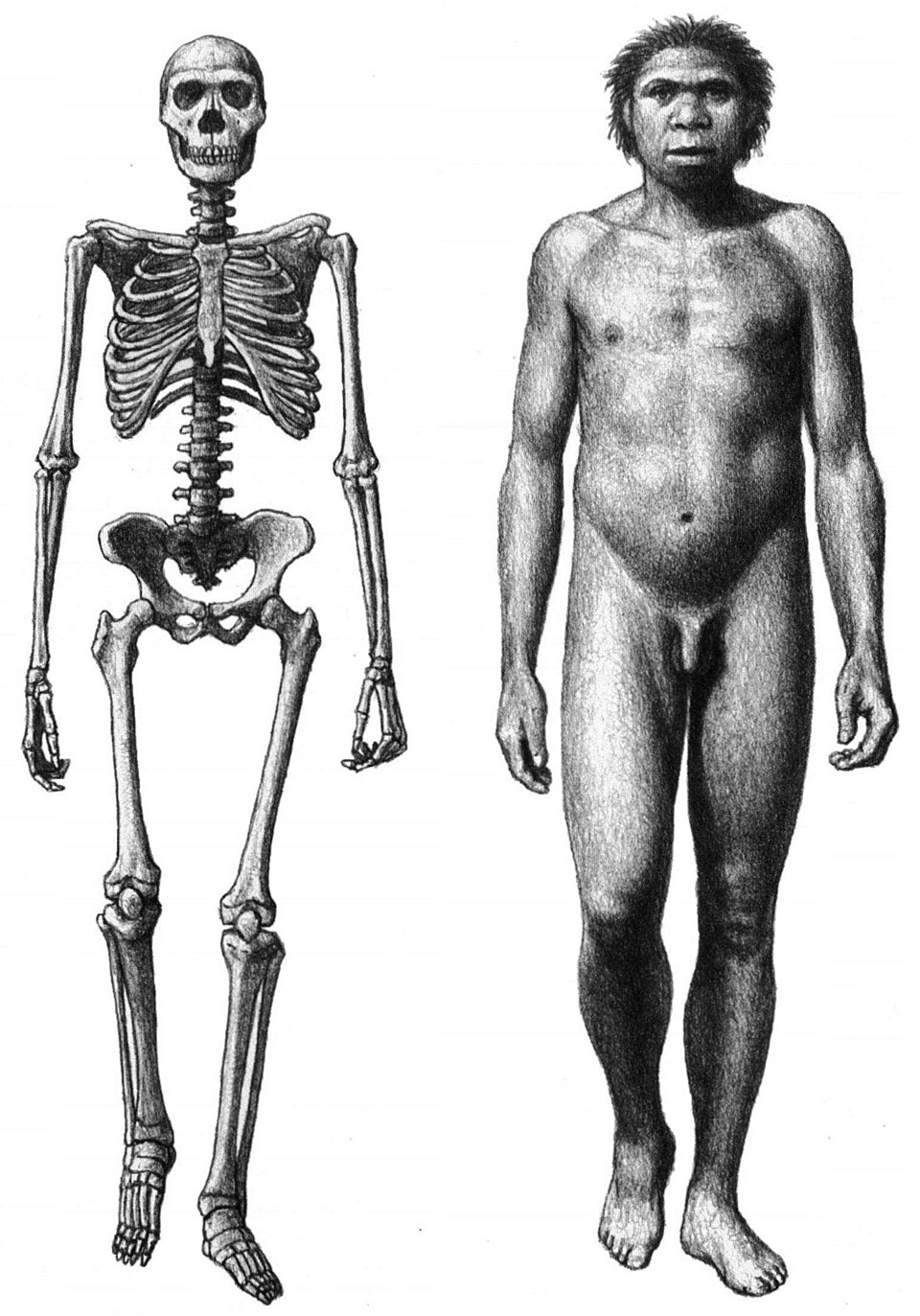
Wrangham argued that this transition should be detectable in the fossil record at the point where a hominin species first shows the anatomical signatures of a cooked-food diet: smaller teeth, reduced jaw musculature, a smaller gut (inferred from torso proportions), and a larger brain. He identified Homo erectus, which appeared approximately 1.9 million years ago in East Africa, as the species that best fits this profile.1, 2 The cooking hypothesis therefore predicts that hominins had achieved some degree of fire control by the time of H. erectus, a claim that has proven to be the hypothesis's most controversial element.
Caloric yield and the energetics of cooking
The empirical foundation of the cooking hypothesis rests on a substantial body of nutritional science demonstrating that thermal processing increases the usable energy in food. The mechanisms differ for the three major macronutrients — carbohydrates, proteins, and lipids — but the net effect is consistently positive.3
{kind=link}
For starch-rich plant foods, the primary mechanism is gelatinization. Raw starch exists as tightly packed semicrystalline granules that resist enzymatic attack in the human small intestine. Much of the starch in raw tubers, roots, and seeds passes through the small intestine undigested and is fermented by bacteria in the colon, a process that yields substantially less metabolizable energy than enzymatic digestion in the small intestine.3 Heating starch in the presence of water causes the granules to swell and lose their crystalline structure, exposing the amylose and amylopectin chains to salivary and pancreatic amylase. Rachel Carmody and Wrangham reviewed the experimental literature in 2009 and documented that cooking raised the proportion of starch digested in the small intestine by 30% to over 100% across five domesticated plant species (oat, wheat, plantain, green banana, and potato), with a median increase in net energy yield of approximately 30%.3 For wild plant foods with higher proportions of resistant starch, the gains from cooking are likely to be at the upper end of this range or higher.
For protein-rich foods, cooking denatures the three-dimensional structure of proteins, unfolding them and making peptide bonds more accessible to digestive proteases. Denatured proteins are hydrolyzed more rapidly and completely in the stomach and small intestine than native proteins, reducing the energetic cost of digestion (diet-induced thermogenesis) and increasing the proportion of amino acids absorbed.3, 4 Cooking also kills parasites and pathogens in meat, reducing the risk of foodborne illness — a benefit that, while not directly caloric, would have had significant fitness consequences for early hominins.
In 2011, Carmody, Weintraub, and Wrangham provided the first controlled experimental evidence that these nutritional effects translate into measurable differences in body mass in a model organism.4 Using mice fed isocaloric diets of meat (lean beef) or starch-rich tubers (sweet potato) in raw or cooked form, they found that mice fed cooked food gained significantly more body mass than mice fed raw food, even when total food intake and activity levels were equivalent. The effect was observed for both meat and tubers, and cooking produced greater gains than pounding (a nonthermal processing technique), demonstrating that heat itself, not merely mechanical softening, was the critical variable.4 These results provided direct experimental support for the proposition that cooking increases net energy gain from food in ways that cannot be captured by traditional proximate analysis, which measures gross energy content rather than metabolizable energy after digestion.
Wrangham and Carmody synthesized these findings in a 2010 review, arguing that the energetic consequences of cooking are sufficiently large to have constituted a decisive selective advantage for any hominin population that achieved reliable fire control.5 An increase of even 20–30% in net energy gain from a staple food source would, over evolutionary time, provide the caloric margin necessary to sustain a substantially larger brain without requiring proportional increases in foraging time or food intake.5
Anatomical predictions and the fossil record
A central strength of the cooking hypothesis is that it generates specific, testable predictions about the anatomical consequences of a shift to cooked food, and these predictions can be evaluated against the hominin fossil record. Wrangham identified four principal anatomical signatures that should accompany the adoption of a cooked diet: reduced tooth size, diminished jaw musculature, a shorter gastrointestinal tract, and an enlarged brain.1, 6
{kind=link}
The dental evidence is consistent with the hypothesis. Homo erectus shows a marked reduction in both molar size and enamel thickness compared with its predecessors in the genus Homo and the australopiths.15 Katherine Zink and Daniel Lieberman demonstrated experimentally in 2016 that the combination of simple Lower Palaeolithic food-processing techniques (slicing with stone flakes) and the inclusion of meat in the diet would have reduced the number of chewing cycles per year by approximately 17% and the total chewing force by approximately 26%, relative to a diet of raw plant foods alone.15 Adding cooking to this baseline would have produced even greater reductions in masticatory demand, providing a plausible selective mechanism for the gracilization of the jaws and teeth documented in the fossil record of early Homo.15
Chris Organ and colleagues applied Bayesian phylogenetic methods to comparative data on feeding time in 2011 and found that modern humans spend an order of magnitude less time feeding than predicted for a primate of our body mass — approximately 4.7% of daily activity time, compared with 48% for chimpanzees and other great apes.17 Extrapolating backward through the hominin phylogeny, they estimated that a significant reduction in feeding time, consistent with the adoption of food processing, occurred at or near the origin of Homo erectus. Molar sizes in H. erectus, H. neanderthalensis, and H. sapiens all fell outside the 99% credible interval for expected values based on phylogeny and body mass alone, suggesting that multiple species in the Homo lineage had adapted to softer, more processed diets.17
The gut cannot be directly observed in the fossil record, but its size can be inferred from the dimensions of the rib cage and pelvis, which constrain the volume available for abdominal organs. The torso of Homo erectus, as reconstructed from specimens such as the Nariokotome Boy (KNM-WT 15000), is narrower and more barrel-shaped than the funnel-shaped torso typical of australopiths, consistent with a reduced abdominal volume and a shorter gastrointestinal tract.1, 5 This morphological change aligns with the prediction of the expensive tissue hypothesis that the gut shrank as the brain expanded, and the cooking hypothesis provides a specific mechanism — thermal food processing — that would have made a smaller gut viable by increasing the digestibility of food before it entered the body.1, 16
Archaeological evidence of fire
The archaeological record of early fire use is fragmentary, contested, and central to the debate over the cooking hypothesis. The difficulty lies in distinguishing anthropogenic fire from natural wildfire in the geological and sedimentary record, and in the poor preservation potential of ephemeral hearths and campfires in open-air settings where most early hominins lived.6, 10

{kind=link}
The earliest widely accepted evidence of fire in an archaeological context comes from Wonderwerk Cave in the Northern Cape province of South Africa. In 2012, Francesco Berna and colleagues published a microstratigraphic analysis of Acheulean-bearing deposits approximately 30 meters inside the cave, dated to roughly 1.0 million years ago.7 Using Fourier transform infrared microspectroscopy and micromorphological analysis of intact sediments, they identified well-preserved ashed plant material and fragments of burned bone deposited in situ on discrete surfaces within the cave, ruling out natural wildfire or post-depositional intrusion as explanations.7 The angularity and preservation of the burned fragments indicated that combustion had occurred locally, close to the find spot, consistent with intentional fire use by the Acheulean tool-makers who occupied the cave.
At Gesher Benot Ya'aqov, an open-air Acheulean site on the shores of a paleo-lake in the northern Jordan Valley of Israel, Naama Goren-Inbar and colleagues documented clusters of burned flint, seeds, and wood in deposits dated to approximately 790,000 years ago.8 The spatial distribution of burned materials in discrete concentrations, rather than as a uniform scatter that might indicate wildfire, was interpreted as evidence for localized hearths.8 Nira Alperson-Afil's subsequent spatial analysis of burned and unburned microartifacts across eight archaeological horizons at the site demonstrated that the pattern of burning was consistent across the stratigraphic sequence, suggesting not a single isolated fire event but continual, repeated fire use over a sustained period of occupation.9
At Qesem Cave in central Israel, Ruth Shahack-Gross and colleagues identified a large central hearth, approximately four square meters in area, with superimposed combustion layers dated to roughly 300,000 years ago.13 The hearth was associated with abundant butchered animal remains and dense concentrations of flint tools, indicating that fire-centered food processing was a habitual and spatially organized activity among the cave's inhabitants.13 Ron Shimelmitz and colleagues analyzed burned flint from a 16-meter-deep stratigraphic sequence at Tabun Cave, also in Israel, and proposed that the transition from sporadic to habitual fire use occurred approximately 350,000 years ago, marking a threshold after which fire became a regular component of hominin technological repertoires across the eastern Mediterranean.12
Wil Roebroeks and Paola Villa's 2011 review of the European evidence reached a similar conclusion: across the continent, convincing evidence of habitual fire use appears only from approximately 300,000–400,000 years ago onward, roughly coinciding with the Middle Pleistocene populations classified as Homo heidelbergensis or early Neanderthals.10 Earlier European sites, including those occupied by H. erectus-grade hominins at Dmanisi (1.77 Ma) and Atapuerca (1.2 Ma), have not yielded unambiguous fire evidence, suggesting that the first hominins to colonize northern latitudes did so without habitual fire control.10
More contested evidence comes from older sites. At Koobi Fora in Kenya, Sarah Hlubik and colleagues have documented thermally altered sediments and artifacts at the FxJj20 site complex, dated to approximately 1.5 million years ago.11 Fourier transform infrared spectrometry revealed patterns of heating consistent with localized combustion rather than natural bushfire, and the spatial association of burned materials with stone tools and faunal remains suggested anthropogenic fire use.11 However, the interpretation of these early sites remains debated, and few researchers consider the evidence before 1 million years ago to be conclusive.
Timeline of archaeological fire evidence7, 8, 10, 11, 12, 13
The evidentiary gap
The most persistent and forceful criticism of the cooking hypothesis concerns the temporal mismatch between its predictions and the archaeological evidence. The hypothesis predicts that cooking should have been adopted at or near the origin of Homo erectus, approximately 1.9 million years ago, to account for the anatomical changes visible in the fossil record at that transition.1, 6 Yet the earliest secure evidence for controlled fire use, at Wonderwerk Cave, dates to approximately 1.0 million years ago, and the first evidence for truly habitual, routine fire use does not appear until 300,000–400,000 years ago.7, 10, 12 This leaves a gap of nearly a million years between the predicted onset of cooking and the earliest uncontested fire evidence, and a gap of approximately 1.5 million years before habitual fire use is firmly documented.
Wrangham has consistently argued that this gap reflects preservation bias rather than genuine absence.1, 6 Open-air fires leave minimal durable traces in the sedimentary record: ash disperses, charcoal degrades, and heat-altered sediments are difficult to distinguish from natural processes without detailed microstratigraphic and spectroscopic analysis of the kind that has only recently been applied systematically to Palaeolithic sites.6 Cave sites, which are far more likely to preserve combustion evidence, are rare in the East African Rift Valley where H. erectus first appeared, and the oldest well-preserved cave deposits in Africa with clear occupation layers tend to postdate 1 million years ago. In a 2017 review, Wrangham argued that the absence of evidence is not evidence of absence, and that the anatomical evidence from H. erectus itself constitutes a form of indirect evidence for cooking that should be weighed alongside the archaeological fire record.6
Critics have responded that the argument from preservation bias, while logically possible, risks making the hypothesis unfalsifiable: if any absence of fire evidence can be attributed to poor preservation, the hypothesis cannot in principle be contradicted by negative findings.10, 18 Nicolas Rolland, in a 2004 review of the Middle Pleistocene record, argued that the pattern of fire evidence across Eurasia is most consistent with a punctuated model in which domestic fire became a regular feature of hominin life only from approximately 400,000–350,000 years ago onward, driven by the cognitive and technological advances of Middle Pleistocene populations rather than by earlier hominins.18 Roebroeks and Villa similarly concluded that the European record shows no convincing evidence of fire control before the late Lower Palaeolithic, and that early hominins in northern latitudes survived without habitual fire for hundreds of thousands of years, suggesting that fire was not yet an obligate component of their adaptive strategy.10
Wrangham's counter-argument points to a distinction between fire production and fire maintenance. Early hominins need not have been able to create fire from scratch; they could have captured and maintained naturally occurring fires from lightning strikes, volcanic activity, or spontaneous combustion of organic material, gradually learning to tend and transport embers before eventually developing the ability to produce fire independently.5, 6 Under this model, the archaeological signature of early fire use would have been even more ephemeral than that of later, technologically more sophisticated fire management, making its recovery in the archaeological record correspondingly less likely. Whether this argument is sufficient to bridge the evidentiary gap remains a matter of active debate.
Fire as social technology
The cooking hypothesis extends beyond nutrition and anatomy to encompass the social consequences of fire control. Wrangham argued that the campfire fundamentally restructured hominin social life by creating a fixed point around which daily activities were organized, by extending the usable hours of the day into the evening, and by establishing a setting for the kinds of sustained social interaction that characterize human societies but are absent in other primates.1, 19
Polly Wiessner's 2014 study of conversation topics among the Ju/'hoansi (!Kung) Bushmen of southern Africa provided empirical support for the social significance of firelight. Wiessner compared 174 daytime and nighttime conversations and found a striking divergence in their content.14 Daytime talk was dominated by economic planning, resource management, and social criticism — the practical business of coordinating group activities and regulating social relationships through gossip. Nighttime firelight talk, by contrast, shifted decisively toward storytelling, singing, ceremonial discussions, and accounts of people in distant social networks.14 Wiessner argued that firelight conversations function to extend the human social imagination beyond the immediate group, evoking higher orders of theory of mind, transmitting cultural institutions, and building the trust and shared understanding that sustain cooperation at the regional level.14
Robin Dunbar and John Gowlett explored the intersection of fire and the social brain hypothesis in a 2014 chapter, arguing that the extension of the active day made possible by firelight would have been critical for maintaining the large social groups predicted by Dunbar's neocortex-group-size relationship.19 In their analysis, the time-budget pressures facing large-brained, large-group-living hominins would have been severe: maintaining social cohesion through grooming, the primary bonding mechanism in other primates, requires time that competes directly with foraging, traveling, and resting. By extending the day and creating a dedicated social space, firelight may have allowed hominins to shift social bonding activities to evening hours, freeing daytime for subsistence tasks and thereby alleviating the time-budget constraint that would otherwise have limited group size.19
The cooking hypothesis also links fire to the evolution of food sharing and the division of labor between the sexes. Wrangham proposed that once cooked food became a central component of the hominin diet, the investment of time and effort required to prepare meals created incentives for pair-bonding and cooperative provisioning: one partner could tend the fire and prepare food while the other foraged or hunted, and the shared meal reinforced the social bond between them.1, 2 This reconstruction remains speculative, as the archaeological record does not directly document the social organization of early hominins, but it illustrates the breadth of the cooking hypothesis's implications for understanding the origins of characteristically human social behavior.
Criticisms and alternative models
Beyond the evidentiary gap in the fire record, the cooking hypothesis has attracted several additional lines of criticism. One prominent challenge concerns the sufficiency of raw food diets. If cooking is as indispensable to human nutrition as Wrangham argues, then pre-cooking hominins should have subsisted on a diet adequate to support moderate brain sizes but not the dramatic encephalization seen in H. erectus. Critics have noted that the transition from australopiths to H. erectus could have been driven by increased meat consumption and simple mechanical food processing (pounding, slicing) rather than by cooking, with cooking adopted later as a supplementary improvement.15 Zink and Lieberman's 2016 experimental work showed that stone-tool-assisted meat processing alone could produce substantial reductions in chewing effort without cooking, suggesting that non-thermal food processing may have been sufficient to drive at least some of the dental and craniofacial changes seen in early Homo.15
A second line of criticism comes from comparative analyses of brain size and fire control. If cooking were the primary driver of encephalization, then the onset of habitual fire use should correlate tightly with periods of rapid brain expansion. However, the most dramatic period of brain growth in the Homo lineage extends from approximately 1.8 to 0.2 million years ago, a period during which fire evidence transitions from absent to contested to routine.10, 20 Herman Pontzer and colleagues' 2016 finding that humans have elevated total energy expenditure relative to other great apes suggested that the metabolic demands of brain expansion were met in part through an absolute increase in energy throughput, not solely through the improved efficiency of food processing that cooking provides.20 This does not refute the cooking hypothesis, but it suggests that cooking was one of several energetic strategies rather than the sole mechanism.
Alternative models for the timing of cooking's adoption have placed its origin considerably later than Wrangham proposes. Rolland's synthesis of the Eurasian record suggested that domestic fire became habitual only around 400,000–350,000 years ago, coinciding with the emergence of Homo heidelbergensis and the late Acheulean, and driven by the cognitive and social complexity of Middle Pleistocene populations rather than by early Homo.18 Under these models, the anatomical changes in H. erectus are attributed primarily to increased meat consumption, improved stone-tool technology, and selection for reduced masticatory apparatus, with cooking entering the picture as a later refinement that accelerated trends already underway.15, 18
Wrangham's 2017 rejoinder to these criticisms reframed the cooking hypothesis in terms of obligation rather than opportunity.6 He argued that the key question is not when hominins first used fire occasionally, which may be archaeologically invisible, but when they became biologically adapted to a cooked diet — that is, when cooked food became so central to their energy budget that they could no longer survive on an exclusively raw diet. The anatomical evidence from H. erectus, in Wrangham's view, indicates that this obligate adaptation had already occurred by 1.9 million years ago, regardless of when the archaeological fire evidence becomes secure.6
Integration with the expensive tissue hypothesis
The cooking hypothesis and the expensive tissue hypothesis are deeply complementary, with the former providing a specific mechanism for the gut-brain trade-off proposed by the latter.1, 16 Aiello and Wheeler's 1995 analysis demonstrated that the human brain is approximately three times larger than expected for a primate of human body mass, while the gastrointestinal tract is only about 60% of its expected size, and that the metabolic savings from the reduced gut approximately offset the metabolic cost of the enlarged brain.16 The expensive tissue hypothesis predicted that encephalization required a shift to higher-quality, more easily digested foods, but it did not specify what that dietary shift was.
Cooking provides a parsimonious answer. By gelatinizing starches, denaturing proteins, and softening fibrous plant material, cooking converts foods that would require extensive gut processing into forms that can be digested rapidly and efficiently in the small intestine.3, 4 A smaller gut is not merely tolerable on a cooked diet; it is sufficient, because the work of breaking down food has been externalized to the fire before the food enters the body. In this framing, cooking is the technological innovation that made the gut-brain trade-off physically possible: the gut could shrink without compromising energy intake, and the freed metabolic resources could be redirected to sustaining a larger brain.1, 5
The integration is not without complications. Subsequent work by Navarrete and colleagues (2011) challenged the strict organ-for-organ trade-off of the expensive tissue hypothesis by finding that, across a broad sample of mammals, the negative correlation between brain size and gut size disappeared when body fat was controlled for.20 And Pontzer and colleagues (2016) demonstrated that humans have genuinely elevated total energy expenditure, suggesting that brain expansion was financed partly through increased energy intake rather than solely through internal reallocation.20 These findings do not invalidate the cooking hypothesis but reframe it: cooking may have contributed to encephalization both by improving the efficiency of digestion (allowing gut reduction) and by increasing total caloric intake (expanding the overall energy budget), with both mechanisms operating simultaneously rather than one exclusively.
Current status
The cooking hypothesis remains one of the most productive and debated frameworks in human evolutionary biology. Its core claims — that cooking dramatically increases the energetic returns from food, that the anatomical changes in Homo erectus are consistent with a transition to cooked food, and that fire profoundly reshaped hominin social life — are broadly supported by experimental, comparative, and ethnographic evidence.1, 3, 4, 14 The hypothesis's central weakness remains the timing question: the gap between the anatomical evidence for a cooked-food adaptation at 1.9 million years ago and the secure archaeological evidence for fire control at 1.0 million years ago has not been closed, and it may not be closable with current archaeological methods given the preservation challenges of detecting early open-air fire use.6, 7
The broader significance of the cooking hypothesis lies in its integration of multiple domains of evidence into a single explanatory framework. By connecting nutritional science, anatomy, archaeology, and social theory, the hypothesis illustrates how a single technological innovation can cascade through an organism's biology and behavior, reshaping teeth, guts, brains, time budgets, and social relationships in a mutually reinforcing process.1, 5, 19 Whether cooking was the initiating cause of these changes at the origin of H. erectus, as Wrangham argues, or a later amplifier of trends set in motion by meat eating and mechanical food processing, as critics contend, the cooking hypothesis has established beyond reasonable doubt that thermal food processing played a major role in making humans the species we are today.
References
Microstratigraphic evidence of in situ fire in the Acheulean strata of Wonderwerk Cave, Northern Cape province, South Africa
Hominin fire use in the Okote member at Koobi Fora, Kenya: new evidence for the old debate
Evidence for the repeated use of a central hearth at Middle Pleistocene (300 ky ago) Qesem Cave, Israel
Impact of meat and Lower Palaeolithic food processing techniques on chewing in humans
The expensive-tissue hypothesis: the brain and the digestive system in human and primate evolution
Was the emergence of home bases and domestic fire a punctuated event? A review of the Middle Pleistocene record in Eurasia