Overview
- The human auditory system is tuned to a frequency range of approximately 2–4 kHz that is critical for speech perception, and fossil evidence from the Middle Pleistocene site of Atapuerca in Spain demonstrates that hominins ancestral to Neanderthals and modern humans had already evolved hearing sensitivity in this speech-relevant range by at least 430,000 years ago.
- The mammalian middle ear evolved from jaw bones of synapsid reptile ancestors, producing the three-ossicle system (malleus, incus, stapes) that provides the impedance matching necessary for efficient airborne sound transmission; this system was inherited by all primates and subsequently fine-tuned in the human lineage.
- Comparative studies of great ape and human hearing show that while chimpanzees hear well at both low and high frequencies, they have a sensitivity dip in the 2–4 kHz range where human hearing is most acute, suggesting that selection specifically enhanced human sensitivity in frequencies critical for distinguishing speech consonants.
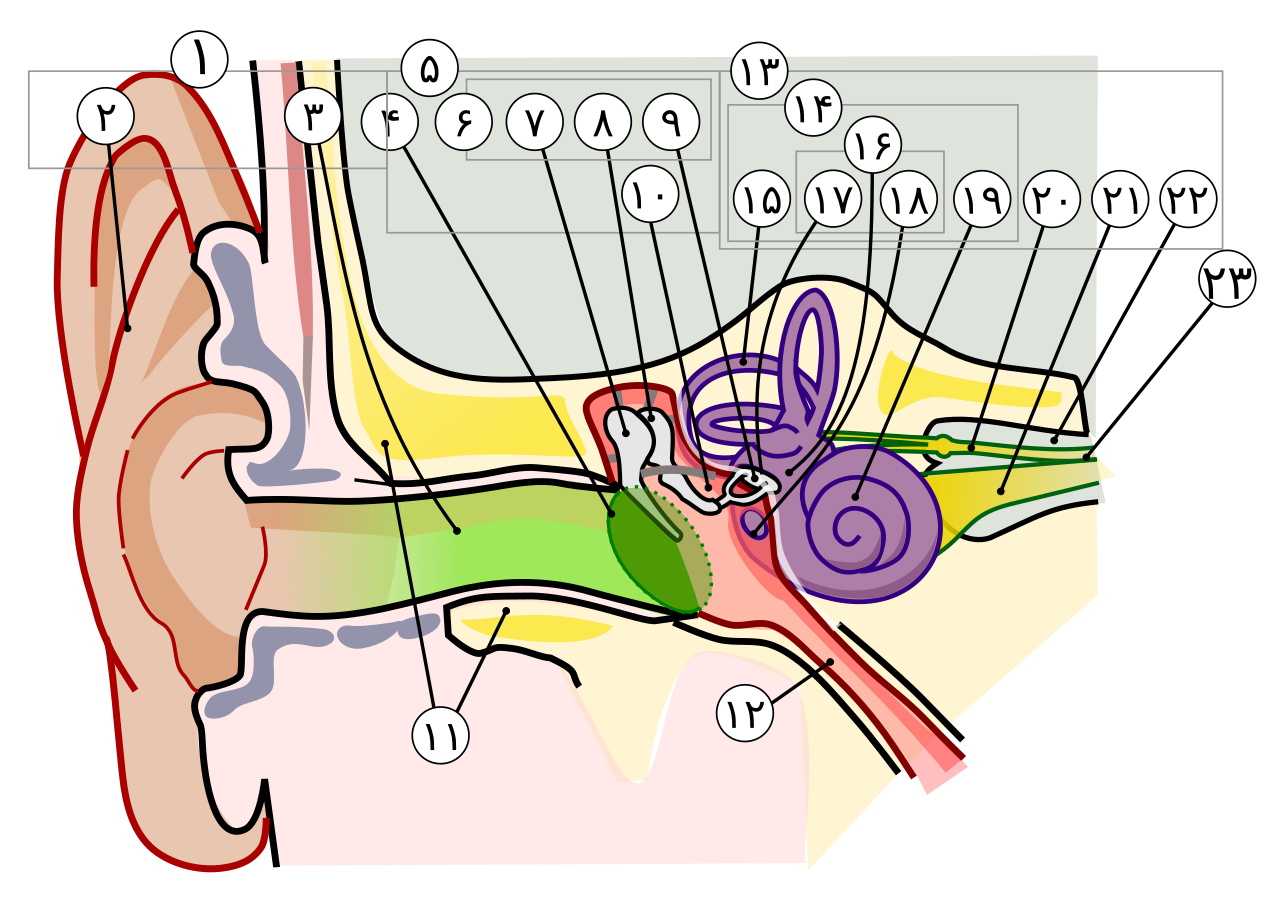
The human auditory system is exquisitely tuned for the perception of speech. While all mammals share the same fundamental ear architecture—an external ear that collects sound, a middle ear that amplifies and transmits vibrations, and a fluid-filled cochlea that transduces mechanical energy into neural signals—the human version of this system exhibits specific refinements in sensitivity, frequency tuning, and cortical processing that distinguish it from the hearing of other primates.6, 16 Tracing the evolutionary history of these refinements is central to understanding when and how spoken language became possible, because effective vocal communication requires not only the ability to produce speech sounds but also the perceptual apparatus to discriminate among them. Fossil evidence from Middle Pleistocene hominins, comparative studies of living primate hearing, and analyses of the anatomy of the middle and inner ear provide converging lines of evidence for how human hearing evolved.1, 2
{kind=link}
The mammalian ear: evolutionary foundations
The three-bone middle ear that characterizes all living mammals—the malleus, incus, and stapes—is itself one of the most remarkable products of vertebrate evolution. In the reptilian ancestors of mammals (the synapsids), the bones that would become the malleus and incus were components of the jaw joint: the articular and quadrate bones, respectively. Over the course of the synapsid-to-mammal transition, documented in a rich fossil series spanning the late Paleozoic and Mesozoic, these bones were progressively reduced in size and decoupled from the jaw, migrating to the middle ear where they were co-opted for sound transmission.5 Zhe-Xi Luo's comprehensive review of this transition documented how the "post-dentary" bones of the synapsid jaw were gradually detached from the mandible through the Mesozoic, becoming the freely vibrating ossicles of the definitive mammalian middle ear by the Cretaceous.5
This three-ossicle system provides the impedance matching necessary for efficient transmission of airborne sound to the fluid-filled cochlea. Sound waves traveling through air encounter a dramatic impedance mismatch at the air-fluid boundary of the inner ear; without the amplification provided by the middle ear's lever system and the area ratio between the large tympanic membrane and the small oval window, approximately 99.9 percent of incoming sound energy would be reflected rather than transmitted.7 The three-ossicle chain overcomes this mismatch far more effectively than the single-ossicle (stapes-only) system of reptiles and amphibians, enabling mammals to hear airborne sounds at lower intensities and across a wider frequency range than most other terrestrial vertebrates.5, 7
Primate hearing and the great ape baseline
Primates as an order have hearing that is broadly typical of mammals, with useful sensitivity spanning roughly 50 Hz to 40–60 kHz depending on the species, and peak sensitivity generally in the range of 1–8 kHz.6 Mark Coleman's comparative analyses of primate audiograms demonstrated that primate hearing sensitivity is influenced by body size, middle ear morphology, and ecological niche, with smaller species tending to hear better at higher frequencies and larger species at lower frequencies.6
The great apes, as the closest living relatives of humans, provide the most relevant baseline for understanding the derived features of human hearing. James Elder's pioneering audiometric work with chimpanzees in the 1930s established that chimpanzee hearing sensitivity is broadly similar to that of humans at both low frequencies (below 1 kHz) and high frequencies (above 8 kHz), but differs notably in the mid-frequency range.3 Subsequent studies confirmed that chimpanzees exhibit a relative dip in sensitivity between approximately 2 and 4 kHz—precisely the frequency range where human hearing is most acute.3, 11
This is a critical observation for understanding speech evolution. The frequencies between 2 and 4 kHz correspond to the formant transitions and consonant cues that carry much of the information content of human speech. Vowels are distinguished primarily by their lower formant frequencies (F1 and F2, typically below 2.5 kHz), but the rapid spectral changes associated with stop consonants, fricatives, and other speech sounds that distinguish words from one another are concentrated in the 2–4 kHz range.13, 16 The enhanced human sensitivity in this band suggests that selection specifically sharpened hearing in the frequencies most relevant for discriminating speech sounds, a refinement not shared by our closest relatives.6, 13
Middle ear anatomy in fossil hominins
The ossicular chain and surrounding middle ear structures are occasionally preserved in hominin fossils, providing direct evidence of auditory capabilities in extinct species. Alexander Stoessel and colleagues conducted high-resolution micro-CT analyses of middle ear ossicles from fossil hominins, modern humans, and great apes, documenting evolutionary changes in ossicular morphology across the lineage.15 Their analyses revealed that the human malleus and incus differ from those of chimpanzees in ways that affect the lever ratio and stiffness of the ossicular chain, modifications that would alter the frequency response of the middle ear and enhance sound transmission in the mid-frequency range relevant to speech.15
Rolf Quam and colleagues extended this approach by developing biomechanical models of the outer and middle ear that can estimate auditory sensitivity from fossil anatomy alone, without requiring a living subject.2 By measuring the dimensions of the ear canal, tympanic membrane, ossicular chain, and oval window in fossil specimens and applying established models of middle ear mechanics, they reconstructed the probable audiograms of extinct hominins. In a 2015 study published in Science Advances, Quam and colleagues applied this method to early hominin fossils from South Africa, including specimens attributed to Australopithecus africanus and Paranthropus robustus, and found that these early hominins had hearing sensitivity patterns intermediate between those of chimpanzees and modern humans.2 Specifically, the South African australopithecines showed slightly enhanced sensitivity in the 1.5–3.5 kHz range compared with chimpanzees, but had not yet achieved the level of sensitivity characteristic of modern humans in the higher speech-relevant frequencies.2
The Atapuerca evidence
The most direct fossil evidence for speech-compatible hearing in an extinct hominin comes from the Sima de los Huesos ("Pit of Bones") site at the Sierra de Atapuerca in northern Spain. This site has yielded the remains of at least 28 individual Middle Pleistocene hominins, dated to approximately 430,000 years ago and attributed to a population ancestral to or closely related to Neanderthals (Homo heidelbergensis sensu lato).1
Ignacio Martínez and colleagues published a landmark analysis in 2004 in which they used CT scanning to measure the middle and outer ear anatomy of five well-preserved temporal bones from Sima de los Huesos and modeled the sound transmission properties of the reconstructed auditory apparatus.1 Their results demonstrated that the Atapuerca hominins had a pattern of sound power transmission through the outer and middle ear that was strikingly similar to that of modern humans and significantly different from that of chimpanzees. In particular, the Atapuerca hominins showed peak sensitivity in the 2–4 kHz range, with a bandwidth of high sensitivity closely matching the modern human pattern and encompassing the frequency range most important for speech perception.1
Martínez and colleagues argued that the human-like hearing pattern observed in the Atapuerca hominins suggests that the auditory prerequisites for speech perception were already in place by at least 430,000 years ago, hundreds of thousands of years before the earliest unambiguous archaeological evidence for symbolic behavior and language in Homo sapiens.1, 14 This does not prove that the Atapuerca hominins had language, but it demonstrates that they had the auditory equipment that, in modern humans, is specifically adapted for processing speech sounds. The finding is consistent with the broader pattern of evidence suggesting that many of the anatomical prerequisites for language evolved incrementally across the Middle Pleistocene rather than appearing suddenly with the origin of modern humans.1, 14
Cochlear evolution and inner ear refinement
The cochlea, the spiral-shaped structure of the inner ear that performs the mechanical-to-neural transduction of sound, is the organ most directly responsible for frequency discrimination. Sound-induced vibrations of the basilar membrane within the cochlea are distributed tonotopically—high frequencies stimulate the base of the cochlea and low frequencies stimulate the apex—and the physical properties of the basilar membrane (width, thickness, stiffness) determine the frequency resolution at each point along its length.9
The human cochlea has approximately 2.5 to 2.75 turns, slightly fewer than the typical chimpanzee cochlea (approximately 2.75 turns), and the basilar membrane dimensions differ in ways that affect frequency tuning.6 Comparative micro-CT analyses of primate cochleae have revealed that humans have a relatively longer basilar membrane in the basal (high-frequency) portion of the cochlea compared with chimpanzees, a feature that may enhance frequency resolution in the mid-to-high frequency range relevant to speech consonant discrimination.6 However, the cochlea is rarely preserved in fossils, and direct evidence of cochlear evolution in hominins remains limited compared with the relatively more accessible middle ear structures.2
Auditory cortex specialization
The perceptual acuity of human hearing depends not only on the peripheral ear structures but also on the specialized processing performed by the auditory cortex. Jon Kaas's comparative studies of auditory cortex organization across primates revealed that humans have a substantially expanded auditory cortex, particularly in the left hemisphere, where speech sounds are preferentially processed.12 The primary auditory cortex (A1) in humans is tonotopically organized, like that of other mammals, but the surrounding belt and parabelt regions are disproportionately enlarged and include areas—notably Wernicke's area and the superior temporal sulcus—that are specialized for processing the temporal and spectral features of speech.12
The left-hemisphere dominance for speech processing, a hallmark of the human brain, has deep evolutionary roots. Functional asymmetries in auditory processing have been documented in other primates, including chimpanzees, which show a left-hemisphere preference for processing species-specific vocalizations, suggesting that the lateralization of auditory processing for communication predates the hominin lineage.12 What appears to be derived in humans is the extent of cortical territory devoted to speech processing and the sophistication of the computations performed, including the ability to parse continuous speech into discrete phonemes, words, and syntactic structures at extremely rapid rates.12, 13
Implications for the timing of spoken language
The fossil and comparative evidence for the evolution of human hearing has direct implications for one of the most debated questions in paleoanthropology: when did spoken language emerge? The Atapuerca evidence demonstrates that by 430,000 years ago, at least one hominin population had evolved hearing sensitivity closely matching the modern human pattern in the speech-frequency range.1 The australopithecine data from Quam and colleagues suggest that the shift toward human-like hearing was already underway by 2–3 million years ago, though it had not yet reached the modern configuration.2
These findings are consistent with a gradual, mosaic model of language evolution in which the anatomical prerequisites—both auditory and vocal—accumulated incrementally over millions of years. Philip Lieberman has argued that the evolution of speech perception and speech production were likely coupled, with improvements in one domain creating selection pressure for improvements in the other in a coevolutionary spiral.13 W. Tecumseh Fitch emphasized that the evolution of language involved not only peripheral adaptations (hearing sensitivity, vocal tract anatomy) but also central neural changes in the capacity for vocal learning, hierarchical sequencing, and intentional communication.16
The auditory evidence alone cannot determine whether the Atapuerca hominins or the South African australopithecines had language in any meaningful sense, because hearing sensitivity is necessary but not sufficient for linguistic communication. However, it does establish a minimum timeline: by the Middle Pleistocene, the ear was ready for speech. The question of when the brain was ready remains one of the most challenging problems in the study of human evolution.1, 2, 16
References
Auditory capacities in Middle Pleistocene humans from the Sierra de Atapuerca in Spain