Overview
- Homo floresiensis was a diminutive hominin species that stood approximately one metre tall with a brain volume of roughly 420 cubic centimetres, discovered in 2003 at Liang Bua cave on the Indonesian island of Flores and dated to approximately 100,000 to 60,000 years ago.
- The hypothesis that the specimens represent pathological modern humans with microcephaly has been extensively tested and rejected through comparative cranial morphology, wrist anatomy, and the discovery of multiple individuals sharing the same distinctive traits.
- The evolutionary origins of H. floresiensis remain debated, with some researchers favouring descent from an early Asian Homo erectus population that underwent island dwarfism, while others argue that cranial and postcranial features point to derivation from a more primitive, pre-erectus hominin that dispersed out of Africa earlier than previously recognized.
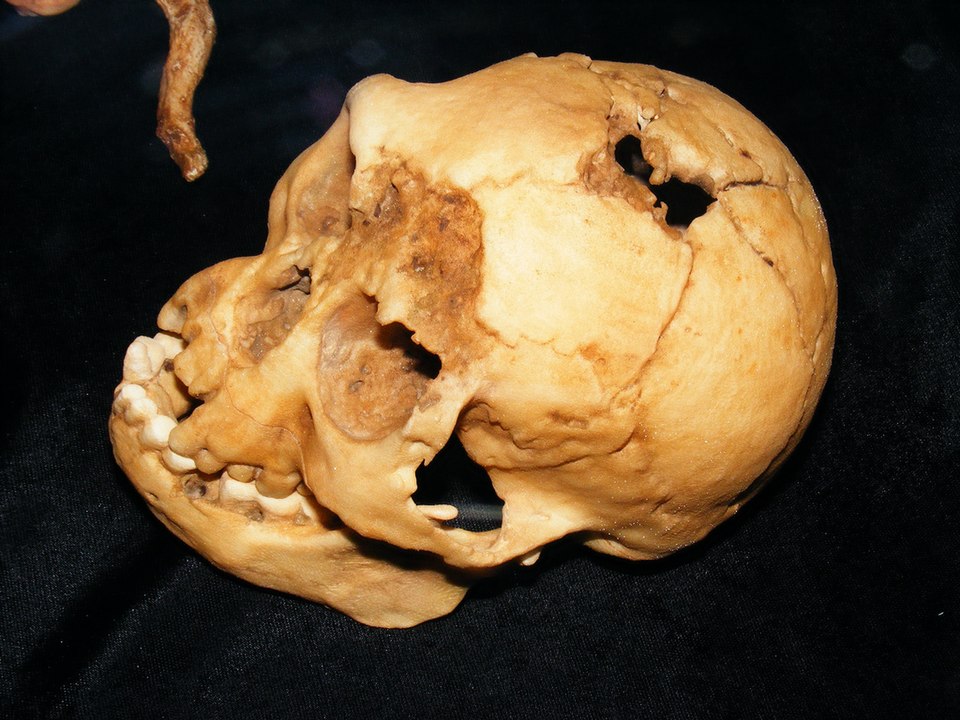
Homo floresiensis is an extinct hominin species known from skeletal remains recovered at Liang Bua, a limestone cave on the Indonesian island of Flores. Announced in 2004 by Peter Brown, Mike Morwood, and colleagues, the species is remarkable for its extraordinarily small body size — approximately 106 centimetres in stature — and a brain volume of roughly 420 cubic centimetres, comparable to that of a chimpanzee and less than a third the average for modern humans.1, 5 The type specimen, designated LB1, is a partial adult skeleton that includes a nearly complete skull, and its discovery immediately challenged prevailing assumptions about the minimum body and brain sizes compatible with stone tool manufacture and complex behaviour in the genus Homo.1, 2 Informally dubbed the "Hobbit" by both the discovery team and the popular press, H. floresiensis has become one of the most debated and consequential hominin discoveries of the twenty-first century.
Discovery at Liang Bua
Liang Bua is a large limestone cave situated in the western part of Flores, one of the Lesser Sunda Islands of eastern Indonesia. Systematic excavation of the cave began in 2001 as a joint project between the Indonesian Centre for Archaeology and the University of New England in Australia, led by Morwood and Raden Soejono. The primary objective was to search for evidence of early modern human (Homo sapiens) migration through the region, but in September 2003, excavators working in Sector VII of the cave uncovered a partial skeleton at a depth of approximately six metres that bore no resemblance to any known modern human population.1, 2
{kind=link}
The skeleton, catalogued as LB1, comprised a skull, mandible, pelvis, partial vertebral column, and elements of both upper and lower limbs. The skull was small but clearly adult, with worn teeth and fused cranial sutures. Brown and colleagues described it as a new species, Homo floresiensis, in October 2004, publishing simultaneously with Morwood and colleagues' description of the archaeological context.1, 2 Subsequent excavations at Liang Bua recovered additional remains representing at least nine more individuals, including a second mandible (LB6) and postcranial elements, all exhibiting the same diminutive proportions and confirming that LB1 was not a singular anomaly.3
Morphology and body proportions
The most striking feature of H. floresiensis is its small size. Based on measurements of the femur and tibia of LB1, stature has been estimated at approximately 106 centimetres and body mass at roughly 25 to 30 kilograms, placing it well below the range of any known adult Homo species and overlapping instead with the body size of Australopithecus afarensis.1 The limb proportions are also unusual: the arms are long relative to the legs, and the humeral-to-femoral ratio resembles that seen in earlier hominins and great apes rather than in modern humans or Homo erectus.1, 8

{kind=link}
The cranium of LB1 has an endocranial volume of approximately 417 to 426 cubic centimetres, depending on the method of measurement, making it the smallest-brained member of the genus Homo by a wide margin.1, 5 Virtual endocasts produced from CT scans of the cranium reveal that despite its small absolute size, the brain of LB1 exhibits a modern-looking frontal lobe morphology, with enlarged Brodmann areas 10 and 47 — regions associated with higher cognitive functions such as planning and initiative — that distinguish it from microcephalic modern humans and suggest a reorganised rather than merely shrunken brain.5
The wrist bones of H. floresiensis are particularly informative. Analysis of the trapezoid, scaphoid, and capitate bones reveals a morphology that is primitive compared to modern humans and Neanderthals, instead resembling the condition seen in great apes and early hominins such as Australopithecus. The trapezoid, for example, is wedge-shaped rather than boot-shaped, a configuration that is derived in modern humans and appears early in the Homo lineage.9 This primitive wrist anatomy is difficult to reconcile with pathological explanations for the small body size and represents strong evidence that H. floresiensis is a distinct species retaining ancestral skeletal features.
The feet of LB1 add another dimension to the morphological picture. At approximately 196 millimetres in length, they are disproportionately large relative to the femur — a ratio unlike that of any known modern human population but consistent with the foot proportions of some African apes and early hominins. The foot lacks a modern longitudinal arch and has a relatively short hallux, features that suggest a biomechanically distinctive gait.12
The dental morphology of H. floresiensis is characterised by relatively large premolars, simplified molar crown morphology, and a suite of traits that do not neatly fit within the known range of variation of any single Homo species. Some dental features resemble those of H. erectus, while others appear more primitive, further complicating phylogenetic placement.21
Chronology and revised dating
The initial dating of the H. floresiensis remains at Liang Bua placed the skeletal material within a broad range of approximately 95,000 to 12,000 years ago, with the youngest deposits containing stone tools potentially as recent as 12,000 years before present. This late survival date generated widespread attention, as it implied that a non-sapiens hominin had persisted on Flores until well into the Holocene, overlapping temporally with the arrival of modern humans in the region.2
In 2016, however, Sutikna and colleagues published a comprehensive revision of the Liang Bua stratigraphy and chronology that significantly altered this picture. New excavations revealed that the original dating had been complicated by an unrecognised erosional unconformity in the cave sediments: younger deposits had been laid down directly against and partly beneath older deposits in a way that created a misleading stratigraphic relationship. Revised uranium-series and radiocarbon dating, combined with luminescence and electron spin resonance ages on associated sediments and fauna, placed the skeletal remains of H. floresiensis between approximately 100,000 and 60,000 years ago, with the associated stone tools extending to approximately 50,000 years ago.4 This revised chronology eliminates any temporal overlap between H. floresiensis and the earliest evidence for modern humans on Flores, though it remains possible that the two species coexisted in the region for a period not yet documented in the archaeological record.
Stone tools and subsistence
The Liang Bua deposits contain a substantial stone tool assemblage associated with H. floresiensis. The tools are made predominantly from volcanic and chert raw materials obtained from the local river system. They are simple in technology, consisting of cores, flakes, and retouched flakes produced by direct hard-hammer percussion, without the prepared-core techniques (such as Levallois) associated with later Homo sapiens and Neanderthal industries.2, 15 Despite their technological simplicity, the tools are consistently produced and show clear evidence of intentional shaping and systematic reduction sequences.16
Faunal remains from the same deposits include bones of the dwarf proboscidean Stegodon florensis insularis, a small-bodied elephant relative endemic to Flores that weighed an estimated 300 to 500 kilograms. Several Stegodon bones bear cut marks and percussion damage consistent with butchery, and some show evidence of burning, strongly suggesting that H. floresiensis hunted or scavenged these animals and processed their carcasses with stone tools.2, 22 The deposits also contain remains of Komodo dragons, giant rats, and various birds, indicating that H. floresiensis exploited a range of the island's fauna. The apparent ability to manufacture stone tools and process large vertebrate prey with a brain one-third the size of a modern human's has profound implications for understanding the relationship between brain size and behavioural complexity in the hominin lineage.2, 5
The pathology debate and its resolution
The announcement of H. floresiensis provoked immediate controversy. Several researchers challenged the interpretation that the Liang Bua specimens represented a new species, arguing instead that LB1 was a pathological modern human. The most prominent alternative hypothesis proposed that LB1 suffered from microcephaly, a condition in which the brain fails to develop to normal size, and that the small body size reflected endemic dwarfism within a modern human population or a growth disorder such as Laron syndrome.6
This pathological hypothesis was subjected to extensive testing over the following decade. Falk and colleagues compared the virtual endocast of LB1 to those of microcephalic humans, normal humans, great apes, and other hominin species and found that the shape of the LB1 endocast differed markedly from microcephalic modern humans, which typically show reduced frontal lobes relative to the rest of the brain. The LB1 endocast instead displayed a pattern of frontal lobe elaboration more consistent with a normally developed, albeit small, brain.5, 7 The discovery of additional individuals at Liang Bua sharing the same small body size and distinctive morphology further undermined the pathology argument, as the probability that multiple unrelated individuals in the same deposit would share the same rare pathological condition is vanishingly small.3
The analysis of the wrist bones by Tocheri and colleagues proved particularly decisive. The primitive trapezoid morphology of H. floresiensis is not a feature associated with any known human pathology; it represents a fundamentally different developmental pathway for the carpal bones that no disease process in modern humans can produce.9 Similarly, the primitive foot morphology described by Jungers and colleagues cannot be attributed to any known modern human pathology and instead points to retention of ancestral hominin features.12 The cladistic analyses by Argue and colleagues, incorporating both cranial and postcranial characters, have consistently placed H. floresiensis as a distinct species within the genus Homo, far outside the range of variation of H. sapiens.8, 19 By the mid-2010s, the pathological hypothesis had been largely abandoned by the palaeoanthropological community.
Island dwarfism
One of the leading explanations for the small body size of H. floresiensis is insular dwarfism, the well-documented evolutionary tendency for large-bodied mammals isolated on islands to evolve reduced body size over generations. This phenomenon, described as part of the broader "island rule," has been observed in numerous mammalian lineages, including elephants, hippos, deer, and bovids, and is thought to result from the combined selective pressures of limited resources, reduced predation, and restricted home ranges on islands.14

{kind=link}
Flores itself provides a striking example of insular dwarfism in a non-hominin lineage: Stegodon florensis insularis, the dwarf proboscidean that coexisted with H. floresiensis, was dramatically reduced in size compared to its mainland ancestors.22 Weston and Lister demonstrated that insular dwarfism in Malagasy hippos was accompanied by disproportionate brain size reduction — the brain shrank faster than the body — and argued that a similar process could explain the unusually small brain of H. floresiensis if the ancestor was a larger-bodied, larger-brained hominin such as H. erectus.13
However, the degree of brain size reduction required to derive the 420 cc brain of H. floresiensis from a typical H. erectus brain of 800 to 1,000 cc is extreme, and Kubo and colleagues demonstrated that the brain-to-body allometric relationship in H. floresiensis falls below the scaling predictions derived from insular dwarfism models applied to H. erectus.17 This finding has led some researchers to propose that the ancestor of H. floresiensis may have been a smaller-bodied, smaller-brained hominin than H. erectus, which would require less extreme dwarfism to reach the observed proportions.
Endocranial volume comparison across hominin species1, 5, 17, 18
Ancestry and phylogenetic position
The evolutionary origin of H. floresiensis remains one of the most debated questions in palaeoanthropology. Two principal hypotheses have been advanced. The first proposes that H. floresiensis descended from Homo erectus, which is known to have been present on the nearby island of Java by at least 1.5 million years ago and which could plausibly have colonised Flores and subsequently undergone insular dwarfism.1, 13 This hypothesis is supported by certain cranial features shared between H. floresiensis and H. erectus, including the overall vault shape and the presence of a supraorbital torus, and by the geographical proximity of Flores to Java.
The second hypothesis, which has gained increasing support, proposes that H. floresiensis descended from a more primitive hominin, possibly one that left Africa before H. erectus. This interpretation is favoured by the suite of primitive postcranial features — the wrist morphology, the foot proportions, the limb ratios, and the shoulder configuration — that are more reminiscent of Australopithecus or early Homo (such as H. habilis) than of H. erectus.8, 9, 12 Cladistic analysis by Argue and colleagues placed H. floresiensis as a sister taxon to H. habilis at the base of the Homo clade, rather than as a descendant of H. erectus, though this result remains contested.19
If H. floresiensis indeed derives from a pre-erectus hominin, the implications for hominin biogeography are profound. It would require that a small-bodied, small-brained hominin dispersed out of Africa and across southern Asia at a much earlier date than the known dispersal of H. erectus, reaching the islands of Southeast Asia without the benefit of the larger body and brain that have traditionally been seen as prerequisites for long-distance dispersal and water crossings.19
The Mata Menge fossils
In 2016, van den Bergh and colleagues reported the discovery of hominin fossils from the site of Mata Menge, an open-air locality in the So'a Basin of central Flores, approximately 70 kilometres east of Liang Bua. The fossils — a fragment of mandible and several isolated teeth — were recovered from deposits dated to approximately 700,000 years ago by a combination of fission-track, argon-argon, and uranium-series methods.10, 20
Critically, the Mata Menge mandible and teeth are as small as or even smaller than the corresponding elements from the Late Pleistocene specimens at Liang Bua. This finding indicates that the diminutive body size characteristic of H. floresiensis had already been achieved by 700,000 years ago, meaning that the dwarfing process must have occurred rapidly after the initial colonisation of Flores.10 Stone tools from the So'a Basin have been dated to approximately one million years ago by fission-track dating, suggesting that hominins first arrived on Flores at or before that time.11, 20 If dwarfism was already complete by 700,000 years ago, the evolutionary transformation from a larger-bodied ancestor to the diminutive Flores hominin occurred within a window of roughly 300,000 years — rapid by evolutionary standards but consistent with observed rates of insular dwarfism in other mammalian lineages.10, 14
The Mata Menge fossils also bear on the ancestry question. Van den Bergh and colleagues noted that the dental morphology of the Mata Menge teeth shares some features with H. erectus from Java, tentatively supporting the H. erectus ancestry hypothesis, though the sample is too small and fragmentary to resolve the debate definitively.10
Implications for hominin dispersal in Island Southeast Asia
The existence of H. floresiensis on Flores has reshaped understanding of hominin dispersal capabilities and the biogeographic history of Island Southeast Asia. Flores lies east of Wallace's Line, the biogeographic boundary that separates the Asian and Australasian faunal realms and that has never been bridged by a land connection, even during periods of maximum glacial sea-level lowering. Reaching Flores from the Asian mainland or from Java required crossing at least two deep-water straits, implying that the ancestors of H. floresiensis either rafted on natural vegetation mats or were capable of some form of deliberate water crossing.11
The stone tool record on Flores extending back to approximately one million years ago demonstrates that hominins reached the island long before the appearance of H. sapiens, and the Mata Menge fossils confirm a hominin presence by at least 700,000 years ago.10, 11, 20 Whether the initial colonisation was an intentional crossing or the result of accidental dispersal (for example, individuals swept out to sea by a tsunami or flood) remains unknown, but the sustained presence on the island over hundreds of thousands of years indicates that a viable breeding population was established and persisted.
The revised dating of the Liang Bua remains to 100,000 to 60,000 years ago raises the question of what caused the disappearance of H. floresiensis. The earliest evidence for modern humans on Flores postdates the last known occurrence of H. floresiensis, and it is possible that the arrival of H. sapiens — with their more sophisticated technology and broader subsistence strategies — played a role in the extinction, whether through direct competition, habitat modification, or the introduction of novel predators and pathogens.4 However, the temporal gap between the disappearance of H. floresiensis and the arrival of modern humans at Liang Bua means that this causal link remains speculative.
The discovery of H. floresiensis, together with the contemporaneous recognition of other archaic hominin populations in Island Southeast Asia — including the Denisovans, known primarily from DNA but with a possible presence in the region — has underscored that the Late Pleistocene was a period of far greater hominin diversity than previously appreciated. Rather than a simple narrative of modern human expansion into an empty landscape, the emerging picture is one of a complex archipelago inhabited by multiple hominin lineages, each adapted to its particular island environment, that may have interacted in ways not yet fully understood.4, 10
References
Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia
Fission-track ages of stone tools and fossils on the east Indonesian island of Flores
Body size, brain size, and sexual dimorphism in Homo naledi from the Dinaledi Chamber
The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters