Overview
- Homo longi ('Dragon Man') was proposed in 2021 as a new human species based on the Harbin cranium, a massive, nearly complete skull from northeastern China with a cranial capacity of approximately 1,420 cubic centimeters and a mosaic of archaic and derived features.
- In June 2025, mitochondrial DNA and protein analysis of dental calculus from the Harbin cranium placed it within the Denisovan clade, suggesting that Homo longi may be synonymous with the Denisovans rather than a separate species.
- The taxonomic debate surrounding the Harbin cranium highlights a broader challenge in paleoanthropology: reconciling morphology-based species designations with molecular phylogenetic evidence, particularly for the diverse array of Middle and Late Pleistocene Homo populations across East Asia.
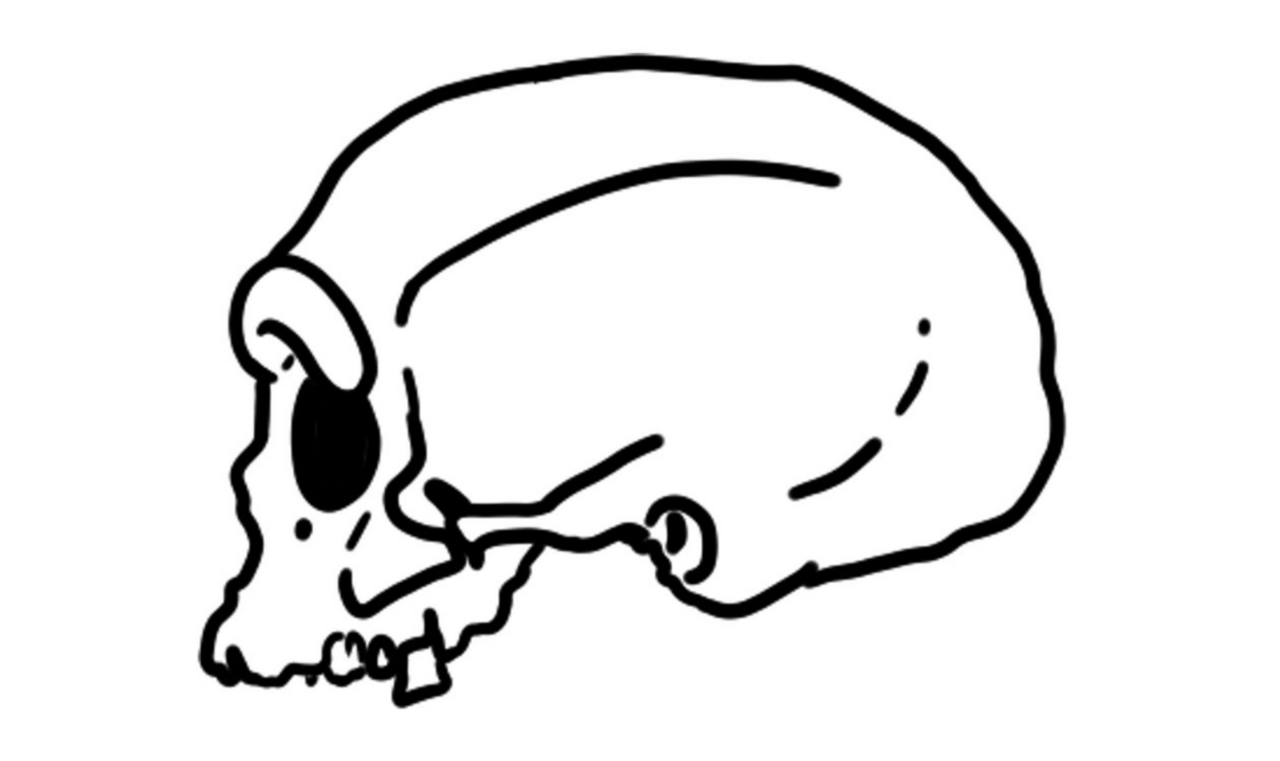
In 2021, a team of Chinese researchers announced the naming of a new human species, Homo longi, based on an exceptionally well-preserved cranium from Harbin, a city in Heilongjiang Province, northeastern China. The species name derives from the Chinese Long Jiang (Dragon River), the historic name of the Heilongjiang (Amur) River, earning the specimen the popular nickname "Dragon Man."1 The Harbin cranium is one of the largest and most complete hominin skulls ever found, combining a massive brow ridge and other archaic features with a cranial capacity of approximately 1,420 cubic centimeters — well within the modern human range.1, 2 However, the species designation was immediately controversial. Many paleoanthropologists suspected that the specimen might represent a Denisovan rather than a genuinely new species, and in June 2025, molecular analyses confirmed that the Harbin cranium falls within the Denisovan clade, casting serious doubt on the validity of Homo longi as an independent taxon.4, 5
{kind=link}
Discovery and provenance
The Harbin cranium has one of the most unusual discovery histories in paleoanthropology. According to the account provided by the family who donated the specimen, the skull was found in 1933 by a laborer working on a bridge over the Songhua River near Harbin, during the Japanese occupation of Manchuria. Rather than surrender the fossil to Japanese authorities, the worker reportedly concealed it in an abandoned well at the family homestead, where it remained undisturbed for more than eighty years. Shortly before his death in 2018, the man told his family about the hidden skull, and they donated it to the Geoscience Museum of Hebei GEO University.1
This circuitous provenance poses significant challenges. Because the cranium was not recovered through controlled excavation, its precise geological context — the sedimentary layer from which it came, its spatial relationship to other fossils or artifacts, and its original depositional environment — is unknown. The research team addressed this problem through geochemical analysis. Rare earth element concentrations and strontium isotope ratios in the cranium bone were compared against those measured in sediment and mammalian fossils from known stratigraphic contexts along the Songhua River. These analyses supported an origin from Middle Pleistocene fluvial deposits in the Harbin area.3 Uranium-series dating of a small bone sample yielded a minimum age of approximately 146,000 years, placing the specimen in the late Middle Pleistocene, while geochemical correlation with dated sediments suggested an age between 138,000 and 309,000 years ago.3
Cranial morphology
The Harbin cranium preserves the face, braincase, and much of the cranial base — an exceptionally complete state of preservation for a Pleistocene hominin fossil. It belonged to a mature male, as indicated by its large size, pronounced muscle markings, and fully fused cranial sutures.1 The specimen presents a striking mosaic of features that do not fit neatly into any single previously defined species.
The braincase is long and low, with a cranial capacity of approximately 1,420 cubic centimeters, comparable to the modern human average and substantially larger than that of typical Homo heidelbergensis or Homo erectus specimens.1 The supraorbital torus (brow ridge) is massive and nearly continuous across the midline, comparable in size to that of archaic Homo species but differing from the double-arched morphology characteristic of Neanderthals. The face is broad and flat, with large, nearly square eye orbits, a wide nasal aperture, and a low, wide infraorbital region. The cheekbones are flat and laterally oriented rather than anteriorly projecting, giving the face a broad, robust appearance distinct from both the midfacial prognathism of Neanderthals and the gracile, vertical face of modern humans.1, 2
The dental arcade is large, with teeth that are substantially bigger than the modern human average but smaller than those of Homo erectus. The maxillary teeth show moderate taurodontism (enlarged pulp cavities), a feature common in archaic Homo but rare in modern populations.1 The cranial base preserves features that distinguish the Harbin cranium from Neanderthals, including a wider and more anteriorly positioned foramen magnum and a differently shaped temporal bone. In overall morphological gestalt, the Harbin cranium most closely resembles other Chinese Middle Pleistocene crania, particularly the Dali cranium from Shaanxi Province, though it is substantially larger.1, 11, 12
The taxonomic proposal
The 2021 publication package comprised three coordinated papers. Ji and colleagues formally described the cranium, erected the new species Homo longi, and provided a differential diagnosis distinguishing it from all other named Homo species.1 Ni and colleagues conducted a phylogenetic analysis using a matrix of 600 cranial and dental characters scored across 95 hominin fossils and found that the Harbin cranium formed a clade with several other Chinese Middle Pleistocene fossils — including the Dali, Jinniushan, and Hualongdong crania — that was the sister group to Homo sapiens, more closely related to modern humans than Neanderthals were.2 If correct, this phylogeny would have profound implications for human evolution, as it would mean that our closest known relatives were not the Neanderthals of western Eurasia but a diverse population of large-brained archaic humans in East Asia.2
The taxonomic designation drew immediate skepticism from many researchers. Several concerns were raised. First, the morphology-based phylogenetic analysis relied on a data matrix that did not include Denisovan specimens — understandably, since the Denisovans at that point were represented primarily by a finger bone, teeth, and the Xiahe mandible, with no cranial material suitable for broad morphological comparison.9 Second, the geographic distribution of Gigantopithecus-associated fossils and the known range of Denisovan DNA in modern populations both pointed to East Asia as a probable center of Denisovan occupation, making the Harbin cranium a plausible candidate for a Denisovan skull.8, 15 Third, the practice of naming a new species from a single specimen with uncertain provenance raised methodological concerns about whether the observed features were genuinely species-diagnostic or simply reflected individual or population-level variation within a known lineage.6
Molecular resolution
The question of whether the Harbin cranium represented a new species or a known one — specifically a Denisovan — could only be resolved through molecular evidence. In June 2025, two landmark studies provided that resolution. Qiaomei Fu and colleagues at the Institute of Vertebrate Paleontology and Paleoanthropology (IVPP) in Beijing successfully extracted mitochondrial DNA and ancient proteins from dental calculus (hardened plaque) adhering to the specimen's teeth.4, 5
The mitochondrial DNA analysis recovered 64 base pairs of the hypervariable region and portions of two coding genes. Phylogenetic placement showed that the Harbin mtDNA fell unambiguously within the Denisovan clade, sharing three derived nucleotide variants found exclusively in known Denisovan specimens (Denisova 3, Denisova 4, and the Baishiya Karst Cave sediment DNA) and sharing none of the derived variants diagnostic of Neanderthals or modern humans.4 The proteomic analysis, conducted on 95 ancient proteins extracted from the same calculus, identified three amino acid variants that matched Denisovan-specific positions and were absent from the Neanderthal and Homo sapiens reference proteomes.5
Together, these results established that the Harbin cranium belonged to a Denisovan individual, making it the first near-complete Denisovan skull and finally revealing what this previously faceless population looked like. The implications for the Homo longi designation are significant: if the Harbin cranium is Denisovan, then the name Homo longi may be synonymous with whatever formal taxonomic name is eventually assigned to the Denisovans — a matter complicated by the fact that the Denisovans were originally defined genetically rather than morphologically and have never been formally described as a species under the International Code of Zoological Nomenclature.6
East Asian archaic diversity
The Harbin cranium does not stand in isolation. Several other Chinese Middle Pleistocene crania share broadly similar morphological features: a large braincase, robust brow ridges, a broad flat face, and a combination of archaic and derived traits that place them in an intermediate position between Homo erectus and modern Homo sapiens. The Dali cranium, discovered in 1978 in Shaanxi Province and dated to approximately 250,000 to 300,000 years ago, shows strong morphological affinities with the Harbin specimen.11, 12 The Hualongdong skull from Anhui Province, dated to approximately 300,000 years ago, presents a similar mosaic of features, with a modern-looking face combined with an archaic braincase.13
These specimens have long defied clean taxonomic assignment. Some researchers have classified them as late Homo erectus, others as Homo heidelbergensis, and still others as "archaic Homo sapiens" — a catch-all category that has gradually fallen out of favor as it obscures rather than clarifies evolutionary relationships.14 The diversity of archaic Homo in the Middle Pleistocene of East Asia may, in light of the 2025 molecular results, reflect the morphological range of one or more Denisovan populations that occupied the region for hundreds of thousands of years.6
The genetic evidence from modern populations is consistent with this interpretation. Studies of Denisovan DNA in present-day humans have identified at least two and possibly three genetically distinct Denisovan populations that interbred with the ancestors of different modern groups.15 The highest levels of Denisovan ancestry are found not in mainland East Asian populations, but in Melanesians, Aboriginal Australians, and the Ayta people of the Philippines, suggesting that at least one Denisovan population occupied Southeast Asia or Island Southeast Asia.15, 16 Whether the morphological diversity evident among Chinese Middle Pleistocene crania corresponds to this genetic diversity — that is, whether the Dali, Hualongdong, and Harbin fossils represent one Denisovan population or several — remains an open question that will require molecular analysis of additional specimens to resolve.6
Implications for hominin taxonomy
The case of Homo longi and the Harbin cranium illustrates a fundamental tension in modern paleoanthropology between morphological and molecular approaches to species identification. Morphology-based phylogenetic analysis of the Harbin cranium placed it as the closest known relative of Homo sapiens — closer than Neanderthals.2 Molecular analysis, by contrast, placed it firmly within the Denisovan clade, which is the sister group to Neanderthals — a position substantially more distant from Homo sapiens than the morphological analysis implied.4, 5 This discordance is not unique to the Harbin case; it echoes longstanding debates about whether morphological similarity necessarily reflects evolutionary relatedness, particularly when convergent evolution, mosaic evolution, or high within-species variability can produce misleading patterns.14
The broader lesson of the Harbin cranium may be that the Middle and Late Pleistocene of Eurasia was home to more morphological and genetic diversity within the genus Homo than traditional taxonomic frameworks have recognized. Neanderthals, Denisovans, Homo sapiens, and possibly other as-yet-unidentified lineages overlapped in time and space, interbreeding where they met and producing populations with complex, mosaic morphologies that resist neat classification into species boxes.8, 10 The Harbin cranium — whatever name it ultimately bears — provides the most complete window yet into the physical appearance of one of these populations, revealing a robust, large-brained hominin that was neither Neanderthal nor modern human but something distinctly its own.1, 4
References
Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage