Overview
- Neanderthals produced sophisticated stone tools using the Levallois technique within the Mousterian tradition, manufactured birch bark pitch adhesives for hafting, and wielded wooden hunting spears, demonstrating advanced planning, material knowledge, and technological versatility.
- Multiple lines of evidence point to symbolic and ritual behavior among Neanderthals, including eagle talon jewelry at Krapina dating to 130,000 years ago, perforated and pigment-stained marine shells in Spain, manganese dioxide pigment collection, and intentional burial of the dead at sites such as La Chapelle-aux-Saints and Shanidar Cave.
- Neanderthals cared for severely injured group members over extended periods, consumed diverse diets ranging from large game to marine shellfish and cooked plants, and possessed the anatomical and genetic prerequisites for spoken language, collectively indicating a behavioral repertoire far more complex than traditionally assumed.
For much of the twentieth century, Neanderthals were regarded as cognitively limited brutes incapable of the symbolic thought, technological innovation, and social complexity attributed to anatomically modern humans. That portrait has been comprehensively dismantled by archaeological discoveries over the past several decades. The material record now documents a species that manufactured sophisticated stone tools using techniques requiring sustained planning and motor expertise, produced synthetic adhesives from birch bark, wielded wooden hunting spears against megafauna, controlled and likely generated fire, adorned themselves with eagle talons and pigment-stained shells, buried their dead in prepared pits, and cared for severely injured group members for years or even decades.1, 9, 12, 19 Collectively, this evidence reveals a behavioural repertoire that, while not identical to that of contemporary Homo sapiens, was far richer and more flexible than traditional models allowed.
Understanding Neanderthal behaviour requires careful evaluation of diverse evidence streams: lithic technology, organic residues, site formation processes, skeletal pathology, ancient DNA, and isotopic chemistry. Each line of evidence is subject to interpretive debate, and the picture that emerges is necessarily one of ongoing refinement rather than settled certainty. What follows is a survey of the major domains of Neanderthal cultural and behavioural life as currently understood from the archaeological and palaeogenomic records.
Stone tool technology
The stone tool traditions most closely associated with Neanderthals are the Mousterian industries of the Middle Palaeolithic, a diverse family of lithic assemblages that spans roughly 300,000 to 30,000 years ago across Europe, the Near East, and parts of Central Asia. The Mousterian was first defined at the rock shelter of Le Moustier in the Dordogne region of France and is characterised by a range of retouched flake tools, including side scrapers, points, and denticulates, produced through several distinct reduction strategies.1 The French prehistorian François Bordes identified five major Mousterian facies — Ferrassie, Quina, Typical, Denticulate, and Mousterian of Acheulean Tradition — that he interpreted as the products of distinct cultural traditions, although subsequent debate has also attributed variability to differences in raw material availability, site function, and reduction intensity.1

{kind=link}
The most technically demanding reduction strategy employed by Neanderthals is the Levallois technique, a method of prepared-core knapping in which the knapper shapes a lithic core into a convex, tortoise-shell-like form before detaching one or more predetermined flakes or points from its surface. The technique requires the knapper to conceptualise the desired end product before beginning work, to establish and maintain specific geometric relationships between the striking platform and the flaking surface, and to manage distal and lateral convexities throughout the reduction sequence.2 These cognitive demands — hierarchical planning, three-dimensional spatial reasoning, and the ability to hold a multi-step operational sequence in working memory — have led many researchers to regard Levallois technology as evidence of advanced technical cognition in Neanderthals.2
In the final millennia of their existence, some Neanderthal populations in western Europe produced the Châtelperronian industry, a transitional technocomplex that combines elements of the preceding Mousterian with features more typical of the Upper Palaeolithic, including blade production, bone tools, and personal ornaments such as pierced animal teeth and ivory pendants. The best-known Châtelperronian assemblage comes from the Grotte du Renne at Arcy-sur-Cure in France, where ornaments and bone awls were found in apparent association with Neanderthal dental remains.23 Whether the Châtelperronian represents independent Neanderthal innovation or the result of contact with incoming modern human populations remains a subject of active scholarly debate, complicated by questions about stratigraphic mixing at key sites. Radiocarbon dating of the Grotte du Renne sequence has revealed some chronological anomalies, leading to continued reassessment of the association between the artefacts and the Neanderthal remains.23
Hafting and adhesive technology
Among the most striking demonstrations of Neanderthal technological sophistication is the production of birch bark pitch, a thermoplastic adhesive used to haft stone tools onto wooden handles. Birch bark pitch is the oldest known synthetic material produced by any hominin, and all confirmed Middle Palaeolithic examples are associated with Neanderthals. The earliest known specimens come from the site of Campitello in Italy, dating to approximately 200,000 years ago, while later finds include pieces from Königsaue in Germany (approximately 80,000 to 43,000 years ago) and a tar-backed flint flake recovered from the Zandmotor beach nourishment near The Hague in the Netherlands, dating to approximately 50,000 years ago.4, 5
The Königsaue specimens are particularly informative. One piece preserves on one surface the impression of a retouched stone tool and on the other the impression of what appears to have been a wooden handle, with a human fingerprint embedded in the pitch, demonstrating that the adhesive was used to join a stone implement to a shaft.5 Recent chemical analysis of the Königsaue tar has identified charcoal inclusions and mineral contaminants consistent with an underground, oxygen-restricted production method rather than the simpler condensation technique that can occur when birch bark is burned in an open fire. This finding has been interpreted as evidence for cumulative cultural learning among Neanderthals, in which technological knowledge was refined and transmitted across generations.5
The Zandmotor tool further extends the evidence. Pyrolysis-gas chromatography-mass spectrometry confirmed the adhesive as birch bark tar, and micro-CT scanning revealed the flint flake partially embedded within the tar mass, indicating deliberate composite tool construction.4 The production of birch bark pitch — regardless of the specific method employed — required Neanderthals to select appropriate raw materials, apply heat in a controlled manner, and combine the resulting adhesive with stone and wood components into a functional composite tool, a sequence that implies substantial procedural knowledge and forward planning.
Hunting strategies and weaponry
The direct evidence for Neanderthal hunting weaponry is dominated by the remarkable wooden spears recovered from the site of Schöningen in Lower Saxony, Germany. First described in 1997, these spruce-wood spears were found in association with stone tools and the butchered remains of at least ten horses. The spears, which range from 1.8 to 2.5 metres in length, were shaped with their centre of gravity placed in the forward third of the shaft — the same design principle used in modern javelins — suggesting they were aerodynamically optimised for throwing.3 Originally dated to approximately 400,000 years ago and attributed to Homo heidelbergensis, revised age estimates based on amino acid geochronology now place the Schöningen spear horizon at approximately 300,000 years ago, with some analyses suggesting an even younger date of approximately 200,000 years ago that would bring them firmly within the Neanderthal temporal range.3
Faunal assemblages from Neanderthal sites across Europe consistently indicate the hunting of large, dangerous game. Species represented include red deer, reindeer, aurochs, bison, wild horse, and woolly rhinoceros, with evidence of systematic butchery including marrow extraction.17, 22 The skeletal trauma patterns observed on Neanderthal remains have been compared to those seen in modern rodeo riders, suggesting frequent close-range encounters with large animals — consistent with ambush or thrusting-spear hunting rather than projectile use from a safe distance.18 This interpretation accords with the general absence of clear projectile technology in the Mousterian toolkit, although the question of whether Neanderthals threw their spears or used them exclusively as thrusting weapons remains debated.
Key evidence for Neanderthal hunting and subsistence3, 17, 20, 21
| Site | Date (ka) | Evidence type | Key finding |
|---|---|---|---|
| Schöningen, Germany | ~300–200 | Wooden spears | Aerodynamically shaped javelin-like weapons with horse remains |
| Spy Cave, Belgium | ~36 | Dental calculus DNA | Woolly rhinoceros and mouflon in diet |
| Bajondillo Cave, Spain | ~150 | Marine shell middens | Earliest known Neanderthal shellfish exploitation |
| Shanidar Cave, Iraq | ~50–65 | Dental calculus starch | Date palms, legumes, and cooked grass seeds |
| El Sidrón, Spain | ~49 | Dental calculus DNA | Mushrooms, pine nuts, moss; no detectable meat |
Use of fire
A comprehensive review of the European archaeological record concluded that the habitual use of fire became a consistent feature of hominin behaviour from approximately 300,000 to 400,000 years ago onward, coinciding with the emergence of Neanderthal populations. From the second half of the Middle Pleistocene, evidence of combustion features — hearths, burned bone, heated sediments, and thermally altered flint — becomes widespread at Neanderthal-occupied sites across Europe.6 The transition from sporadic to habitual fire use was a major behavioural threshold, enabling cooking, warmth in glacial environments, protection from predators, and the heat-treatment of materials.
Whether Neanderthals could actively generate fire or merely maintained fires started by natural ignition sources such as lightning was long uncertain. Two lines of evidence now suggest deliberate fire production. At the site of Pech-de-l'Azé I in southwestern France, Neanderthals collected blocks of manganese dioxide, a mineral that substantially lowers the auto-ignition temperature of wood and accelerates the rate of char combustion when ground to powder and applied to tinder. Compositional analysis showed that the manganese dioxide was selectively sourced from deposits several kilometres from the site, ruling out incidental collection, and experimental combustion tests confirmed its effectiveness as a fire-starting aid.7
Independent evidence comes from microwear analysis of dozens of late Mousterian bifaces from multiple French sites. Microwear traces on the flat surfaces of these tools — polish, striations, and impact marks — are consistent with the repeated percussion of iron pyrite against the flint surface, a technique that generates sparks capable of igniting tinder. The wear patterns match those produced experimentally by striking pyrite against flint in a fire-making context and differ from traces left by other uses such as butchery or woodworking.8 Taken together, the manganese dioxide and pyrite-strike evidence indicates that at least some late Neanderthal populations possessed reliable, intentional fire-making technology.
Symbolic behavior
The question of whether Neanderthals engaged in symbolic behaviour — the use of objects or actions to represent ideas beyond their immediate functional purpose — has been among the most contentious issues in palaeoanthropology. The accumulating evidence now makes it difficult to deny some form of symbolic capacity, although its nature and extent remain debated.
.jpg){kind=link}
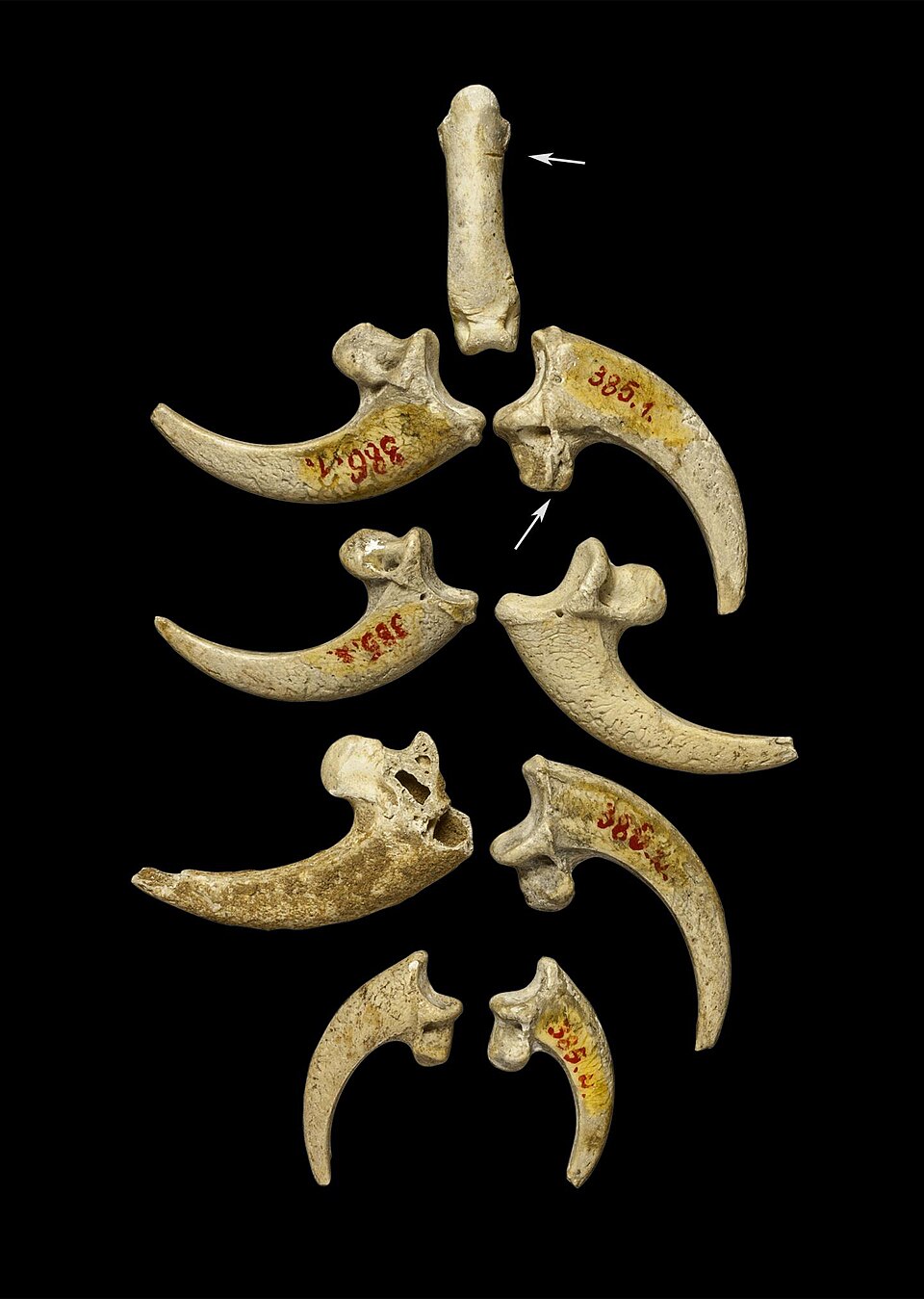
The oldest proposed example of Neanderthal body ornamentation is a set of eight white-tailed eagle (Haliaeetus albicilla) talons recovered from the site of Krapina in Croatia, dating to approximately 130,000 years ago. The talons, which derive from at least three individual eagles, bear cut marks, polishing facets, and small notches at consistent locations along the plantar surface, features interpreted as evidence that the talons were strung together as a necklace or bracelet. The acquisition of talons from multiple raptors, likely requiring live capture, implies planning, patience, and a motivation that is difficult to explain in purely utilitarian terms.9
Additional evidence for ornament use comes from the Iberian Peninsula. At Cueva de los Aviones and Cueva Antón in southeastern Spain, Neanderthal-associated deposits dated to approximately 115,000 years ago yielded perforated marine shells alongside lumps of red and yellow mineral pigments. One Spondylus shell contained residues of a complex pigment mixture composed of a red lepidocrocite base mixed with ground hematite and pyrite fragments, a preparation with no obvious functional utility other than as a cosmetic or decorative substance.10
The use of pigments at Neanderthal sites extends beyond body decoration. While manganese dioxide blocks at Pech-de-l'Azé I appear to have served a pyrotechnological function, the collection and grinding of colourants at other sites may reflect symbolic or aesthetic motivations.7, 10 Perhaps the most dramatic claim for Neanderthal symbolic expression comes from uranium-thorium dating of carbonate crusts overlying cave paintings at three Spanish sites — La Pasiega, Maltravieso, and Ardales — which yielded minimum ages exceeding 64,800 years, predating the arrival of modern humans in Europe by at least 20,000 years and implying Neanderthal authorship. These results, published in 2018, remain controversial, with critics questioning the reliability of the dating methodology and the selection of samples.11
Burial practices
The question of whether Neanderthals intentionally buried their dead has been debated since the early twentieth century. The most thoroughly reinvestigated case is La Chapelle-aux-Saints in southwestern France, where the largely complete skeleton of an elderly Neanderthal was discovered in 1908 in a shallow pit within a small limestone cave. A twelve-year re-excavation programme completed in the 2010s examined the taphonomic context of the original find and concluded that the pit was deliberately dug by other members of the group, that the body was placed within it in a flexed position, and that rapid sedimentary covering protected the skeleton from scavenging and disturbance.12 The excellent preservation of the skeleton, with its delicate elements still articulated, contrasts sharply with the fragmentary and scattered condition of animal bones in the surrounding deposits, reinforcing the conclusion that the burial was intentional rather than the result of natural sedimentary processes.12

{kind=link}
The most famous — and most debated — case is the so-called flower burial at Shanidar Cave in Iraqi Kurdistan. During excavations in the 1950s and 1960s, Ralph Solecki discovered the remains of multiple Neanderthal individuals in the cave, including Shanidar IV, around whom soil samples contained concentrated clumps of pollen from several flowering plant species including yarrow, cornflower, and grape hyacinth. The palynologist Arlette Leroi-Gourhan interpreted these as the remains of bouquets deliberately placed with the body, a conclusion that Solecki popularised in his 1971 book Shanidar: The First Flower People.13, 24
The flower burial hypothesis has been contested on multiple grounds. Critics have noted that the pollen could have been introduced by burrowing rodents or ground-nesting bees, both of which are known to bring plant material into underground chambers. Recent reanalysis of Leroi-Gourhan's original pollen data found that some clumps contained pollen from multiple species — an unlikely occurrence in a naturally deposited flower anther — and identified both ancient and modern burrows made by solitary bees within the Shanidar sediments.24 However, renewed excavations at Shanidar beginning in 2015 uncovered a new articulated Neanderthal skeleton (Shanidar Z) in close proximity to the original flower burial, in a position consistent with deliberate placement, suggesting that the cave was indeed used for repeated funerary deposition regardless of how the pollen arrived.13
Evidence for language capacity
Whether Neanderthals possessed spoken language is a question that cannot be definitively resolved from the archaeological record alone, but two independent lines of evidence suggest that they had the anatomical and genetic prerequisites. The first is the discovery of a virtually complete hyoid bone from the Kebara 2 Neanderthal skeleton, recovered from Kebara Cave on Mount Carmel in Israel and dated to approximately 60,000 years ago. The hyoid is a small, horseshoe-shaped bone in the throat that anchors the muscles of the tongue and larynx and plays a critical role in speech production. The Kebara 2 hyoid is nearly identical in size and morphology to those of modern humans, in contrast to the very different hyoids of chimpanzees and other great apes.15 Subsequent micro-biomechanical analysis using synchrotron-based micro-CT scanning confirmed that the internal architecture and mechanical properties of the Kebara 2 hyoid are also indistinguishable from those of modern human hyoids, suggesting that the position and function of the Neanderthal larynx were comparable to our own.15
The second line of evidence is genetic. In 2007, researchers extracted ancient DNA from Neanderthal skeletal remains from the site of El Sidrón in Spain and determined that Neanderthals shared with modern humans the two amino acid substitutions in the FOXP2 gene that distinguish the human version of this protein from that of chimpanzees. FOXP2 is the only gene for which mutations are known to cause a specific speech and language disorder in humans: individuals with disrupted FOXP2 function exhibit severe impairments in the fine motor control of the orofacial musculature required for fluent speech, as well as broader deficits in grammatical processing.14 The fact that Neanderthals carried the derived human form of FOXP2 indicates that the selective pressures favouring this variant acted before the divergence of the Neanderthal and modern human lineages, more than 300,000 years ago. While the presence of the derived FOXP2 variant does not prove that Neanderthals spoke in the manner of modern humans, it removes one of the proposed genetic barriers to complex vocal communication.14
Diet diversity
The traditional image of Neanderthals as obligate carnivores specialising in large-game hunting has been replaced by a more nuanced picture of dietary flexibility that varied with local ecology. Isotopic analyses of Neanderthal bone collagen, which typically show elevated nitrogen-15 values consistent with a high trophic position, initially supported the meat-dominated model. However, isotopic data reflect protein intake over the final years of life and may underrepresent the contribution of plant foods, which are lower in nitrogen.22
Direct evidence for plant consumption has come from the analysis of microfossils and ancient DNA preserved in dental calculus. Starch grains and phytoliths recovered from the teeth of Neanderthals at Shanidar Cave and Spy Cave in Belgium document the consumption of date palms, legumes, and grass seeds from the tribe Triticeae, some of which had been cooked, as indicated by damage patterns on the starch granules consistent with exposure to water and heat.20 Ancient DNA extracted from the dental calculus of Neanderthals at El Sidrón in Spain revealed a diet that included mushrooms, pine nuts, and forest moss, with no detectable animal DNA, in stark contrast to calculus from Spy Cave Neanderthals whose DNA profile was dominated by woolly rhinoceros and mouflon.17 This geographic variation in diet is consistent with molar macrowear studies showing that Neanderthals in forested Mediterranean environments consumed more hard, abrasive plant foods than those in open steppe-tundra habitats, where meat dominated.22
Marine resources also featured in Neanderthal subsistence. At Bajondillo Cave in southern Spain, shell middens dominated by mussels date to Marine Isotope Stage 6, approximately 150,000 years ago, making this the earliest known evidence of systematic marine shellfish exploitation by Neanderthals and roughly contemporaneous with the earliest shellfish use by modern humans at Pinnacle Point in South Africa. Taphonomic analysis showed that the shells had been broken in a stereotypical manner and many bore thermal alterations consistent with cooking.21
Self-medication and medicinal plant use
Perhaps the most unexpected dimension of Neanderthal behaviour to emerge from recent research is evidence for the deliberate ingestion of plants with medicinal properties. The initial discovery came from the analysis of dental calculus from an individual at El Sidrón in Spain, in which molecular compounds characteristic of yarrow (Achillea millefolium) and chamomile (Matricaria chamomilla) were identified. Both plants are bitter-tasting and possess negligible caloric value, making it unlikely that they were consumed for nutrition. Yarrow has long been used in traditional medicine as an anti-inflammatory and astringent, and chamomile as a digestive aid and mild sedative, raising the possibility that their consumption by Neanderthals reflected an awareness of their therapeutic properties.16
Subsequent ancient DNA analysis of dental calculus from the same El Sidrón population provided further support. One individual who exhibited a dental abscess and carried DNA from the gastrointestinal pathogen Enterocytozoon bieneusi also showed traces of poplar bark, which contains salicylic acid (the active compound in aspirin), and DNA from the mould Penicillium, which naturally produces antibiotic compounds. The co-occurrence of pathological indicators and plant-derived medicinal compounds in the same individual is suggestive, though not conclusive, of intentional self-treatment.17
The self-medication hypothesis remains contested. Critics have argued that the ingestion of bitter or medicinal plants could have been incidental, that the molecular evidence does not distinguish between dietary and therapeutic consumption, and that attributing medical knowledge to Neanderthals requires a level of inferential reasoning about internal bodily states for which the archaeological evidence is inherently ambiguous.16 Nevertheless, the consistency of the pattern across multiple individuals and analytical methods has made the hypothesis a serious subject of ongoing investigation.
Social care for the injured
Some of the most compelling evidence for the social complexity of Neanderthal groups comes from individuals who survived severe injuries that would have rendered them incapable of independent survival. The paradigmatic case is Shanidar 1, a Neanderthal male from Shanidar Cave who survived into his forties — an advanced age for a Pleistocene hominin — despite a catalogue of debilitating pathologies. He had sustained a crushing blow to the left side of his face that may have blinded his left eye, a fracture of the right arm that resulted in its withering and eventual amputation above the elbow, injuries to his right knee and foot that produced a pronounced limp, and degenerative bony growths in both ear canals that would have caused near-total deafness.18
The survival of an individual so severely impaired in a Pleistocene environment teeming with large carnivores implies sustained provisioning and protection by other members of his social group. The loss of his right arm would have severely limited his ability to participate in tool manufacture, hunting, or many routine tasks, while his deafness would have made him unable to detect approaching predators. His survival for years or decades after sustaining these injuries is difficult to explain without invoking some form of social care.18, 19
Shanidar 1 is not an isolated case. Across the Neanderthal fossil record, numerous individuals exhibit healed fractures, amputations, severe dental disease, and degenerative conditions that would have required extended periods of reduced activity and reliance on group support. A systematic review of Neanderthal healthcare documented cases of apparent wound treatment, reduction of activity during recovery, and long-term provisioning of food to impaired individuals.19 Whether this care was motivated by compassion, kinship obligation, or pragmatic calculation about the value of experienced group members is impossible to determine from the skeletal evidence alone, but the pattern clearly indicates that Neanderthal social groups were capable of sustaining non-productive individuals over extended periods — a behaviour that requires foresight, cooperation, and a degree of social cohesion that goes well beyond the capacity of any non-human primate.19
References
Middle Paleolithic complex technology and a Neandertal tar-backed tool from the Dutch North Sea
Production method of the Königsaue birch tar documents cumulative culture in Neanderthals
Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus
Neanderthal behaviour, diet, and disease inferred from ancient DNA in dental calculus
Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium)
Chronology of the Grotte du Renne (France) and implications for the context of ornaments and human remains within the Châtelperronian
Shanidar et ses fleurs? Reflections on the palynology of the Neanderthal ‘Flower Burial’ hypothesis