Overview
- The Taung Child is a fossilised juvenile cranium and natural endocast from the Buxton–Norlim Limeworks in South Africa, described by Raymond Dart in Nature on 7 February 1925 as the type specimen of Australopithecus africanus—the first early hominin recovered from the African continent.
- Dart's interpretation of the small-brained, small-toothed, anteriorly placed foramen magnum specimen as a bipedal human ancestor was rejected for more than two decades by senior British anatomists who favoured the now-exposed Piltdown forgery, and was vindicated only after Robert Broom's discoveries of adult australopiths at Sterkfontein in the late 1930s and 1940s.
- Modern work places the child's age at death between roughly 3.7 and 4 years, dates the deposit close to the Plio–Pleistocene boundary at about 2.6–2.8 million years, and explains the cranium's puncture marks and orbital damage as evidence that the child was killed and partially eaten by a large bird of prey, most likely a crowned eagle.
The Taung Child is a fossilised juvenile cranium, partial face, and natural braincase endocast recovered from the Buxton–Norlim Limeworks of South Africa's North West Province in 1924. The Australian-born anatomist Raymond Dart described the specimen the following year in Nature, designating it the type specimen of a new genus and species, Australopithecus africanus—the "southern ape from Africa."1, 2 It was the first early hominin fossil ever recovered from the African continent and, more than seventy years before molecular evidence settled the matter, the first physical confirmation of Charles Darwin's 1871 prediction that humanity's deep ancestry would be traced to Africa.2
The specimen's importance is out of all proportion to its size. It is the cranium of a child who died at roughly four years of age, with first molars only just erupting through the gum, in a wooded landscape close to the Plio–Pleistocene boundary about 2.6 to 2.8 million years ago.3, 18 Yet a single small face, a partial mandible, and a natural endocast preserved enough information to anchor a new species, overturn the European bias of early-twentieth-century palaeoanthropology, and—through more than a century of reanalysis—feed continuing debate about hominin brain evolution, dental ontogeny, taphonomy, and the depositional environments of South Africa's earliest australopiths.2, 10, 16
Discovery at the Buxton–Norlim Limeworks
The chain of events leading to the recognition of the Taung Child began not with Dart but with one of his students. Josephine Salmons, the first female student of anatomy under Dart at the University of the Witwatersrand, noticed a fossilised baboon cranium displayed on the mantelpiece at the home of the Izod family, whose connections in the Northern Lime Company gave them access to fossils recovered from the Buxton quarries (later known as the Buxton–Norlim Limeworks) near the village of Taung in what is today South Africa's North West Province.2, 21 Recognising the specimen's potential significance, Salmons brought it to Dart's attention. Dart in turn asked the consulting geologist Robert Young to forward any unusual fossils that the limeworkers should encounter while quarrying the brecciated tufa and calcrete deposits.2
In November 1924 two crates of breccia and rubble arrived at Dart's home in Johannesburg as he was dressing to serve as best man at a friend's wedding. Inside the second crate, encased in hard pinkish carbonate matrix, lay an endocranial cast of remarkable detail. Dart immediately recognised the impression as that of a brain markedly larger and more convoluted than any baboon's, and as he later recounted, he could see a face still buried in the surrounding stone that the endocast must once have fitted.2, 21 Over the following weeks Dart used a knitting needle, a small hammer, and his wife's hatpins to chip the matrix away from the bone, freeing the face on 23 December 1924—an event later memorialised as the moment Africa's claim to the human story was first physically demonstrated.3, 21
{kind=link}
Forty days after first opening the crate, Dart had drafted a manuscript describing the specimen. It was telegraphed to Nature in London and appeared in the issue of 7 February 1925 under the title "Australopithecus africanus: The Man-Ape of South Africa."1 Dart was twenty-nine years old.
The 1925 Nature paper
Dart's Nature paper is brief—four pages, illustrated with line drawings of the specimen in lateral, basal, and frontal view—but it is one of the most consequential short publications in the history of palaeoanthropology.1, 2 Dart erected a new genus and species, Australopithecus africanus ("southern ape from Africa," combining the Latin australis with the Greek pithekos), and assigned it to a new family, the Homo-simiadae, intermediate between living anthropoid apes and humans.1, 5
Several anatomical observations carried the argument. The face was less prognathic than that of any living African ape of comparable age, with a relatively shallow muzzle and small canines lacking the projecting, dagger-like form characteristic of male chimpanzees and gorillas. The dentition was hominin in cusp pattern, occlusion, and degree of crowding, and the deciduous and just-erupting permanent molars showed wear patterns inconsistent with extant African apes.1, 2 The cranial base preserved a foramen magnum—the opening through which the spinal cord exits the skull—positioned more anteriorly and inferiorly than in any extant great ape, an arrangement consistent with a vertebral column held beneath, rather than behind, the skull. Dart inferred from this geometry that the animal in life had habitually carried its head erect and walked on two legs, freeing the hands from a primary role in locomotion.1, 2, 24
To these skeletal arguments Dart added a neuroanatomical claim. The natural endocast preserved the impression of the lateral surface of the right cerebral hemisphere, including a number of sulci. Dart identified what he interpreted as a posteriorly placed lunate sulcus, a fissure that in living apes lies relatively far forward and that he took to mark the boundary of the visual cortex. A more posterior position would imply expansion of the parietal association areas at the expense of primary visual cortex—a humanlike condition.1, 13 Combined, the dental, basicranial, and endocranial observations led Dart to conclude that the Taung Child belonged to "an extinct race of apes intermediate between living anthropoids and man" and that Africa, not Europe or Asia, must therefore be the cradle of humanity.1, 2
The British establishment's rejection
The reception was almost uniformly hostile. In the very next issue of Nature, four senior figures in British anatomy and palaeoanthropology—Sir Arthur Keith, Grafton Elliot Smith, Sir Arthur Smith Woodward, and W. L. H. Duckworth—published comments on Dart's announcement, three of them dismissing or sharply qualifying his conclusions.2, 26 Keith, the dominant English voice in human-origins research, argued that Taung's apparent humanlike features were artefacts of immaturity. The skulls of infant chimpanzees and gorillas, he pointed out, look superficially more humanlike than those of adults of the same species: short faces, relatively large braincases, modest canines. Without the corresponding adult skull, Keith insisted, no claim of intermediacy could stand.2, 26
Elliot Smith—the very anatomist under whom Dart had trained at University College London—was politer but no more enthusiastic, treating Taung as an interesting fossil ape unlikely to bear directly on the human lineage.2, 21 Smith Woodward, the British Museum palaeontologist who had described and championed the Piltdown remains, regarded the specimen as a juvenile chimpanzee or gorilla, and the popular press took its cue from the senior figures: cartoons and editorials in 1925 routinely treated the "missing link from Africa" as the overreach of a provincial colonial.2, 26
The hostility was compounded by other features of Dart's paper that grated on professional sensibilities. The species name was a Latin–Greek hybrid, regarded as philologically barbarous; the manuscript had been rushed into print in Nature rather than placed before the Royal Society or the Zoological Society; and Dart, working from Johannesburg, had not first circulated the specimen to the senior authorities of the field. The combination of unorthodox conclusions, unorthodox process, and unorthodox terminology made rejection easy.2, 21, 26
The Piltdown distraction
British resistance to the Taung Child cannot be understood apart from the Piltdown remains, fragments of a cranium and mandible recovered between 1908 and 1915 from gravel pits at Piltdown in Sussex, England, and announced as Eoanthropus dawsoni, the "dawn man." Piltdown combined a large, modern-looking braincase with a chimpanzee-like jaw, and it appeared to confirm an influential prior expectation: that the brain had expanded first, and that other humanlike features had followed in its train.2, 26
The Taung Child reversed this scheme. Its braincase was small—comparable in absolute volume to a juvenile chimpanzee—while its dentition and basicranium were already markedly hominin. Accepting Taung therefore required rejecting the model on which Piltdown's significance depended, and the senior figures who had committed their reputations to Eoanthropus were the very figures whose endorsement Dart needed.2, 26 Even before Piltdown was exposed as a deliberate forgery in 1953—the mandible was that of an orangutan, the cranium a stained modern human, the assemblage chemically and morphologically incoherent—Dart had noted in his 1925 paper that Piltdown's mandibular symphysis "scarcely differs from the anthropoids," a remark that reads in retrospect as quietly prescient.1, 2
The Piltdown forgery's persistence delayed acceptance of the Taung Child by a full generation. While the British establishment defended a fossil that did not exist, Dart largely withdrew from active palaeoanthropology, leaving the defence of Australopithecus africanus to the Scottish-born physician and palaeontologist Robert Broom.2, 21
Sterkfontein and vindication
Broom, then in his early seventies and working out of the Transvaal Museum in Pretoria, took up the search for adult australopiths in 1936. Within months he had recovered fragmentary adult australopith remains from the dolomitic cave deposits at Sterkfontein near Krugersdorp, the first adult specimens of Dart's species ever found.20, 21 In 1938 he announced a second species, Paranthropus robustus, from the nearby Kromdraai site, and in 1947 he and his assistant John Robinson recovered Sts 5, a nearly complete adult cranium from Sterkfontein later popularised under the affectionate label "Mrs. Ples."19, 20 Mrs. Ples confirmed that Australopithecus africanus retained a small braincase and projecting face into adulthood; Keith's "infant chimpanzee" objection collapsed under the weight of the new specimens.19, 20
The endorsement that finally tipped the wider community came in 1947, when the Oxford anatomist Wilfrid Le Gros Clark visited the South African collections, examined the australopith material directly, and reported that the cranial, dental, and mandibular features were consistent with hominin status rather than with any living ape. Within months of Le Gros Clark's report, Sir Arthur Keith publicly conceded the point in a letter to Nature, writing: "I am now convinced...that Prof. Dart was right and that I was wrong."2, 21, 26 The retraction did not appear in the same prominent place as the original dismissal, but it ended professional resistance to the genus that Dart had erected twenty-two years earlier.
Anatomy of the specimen
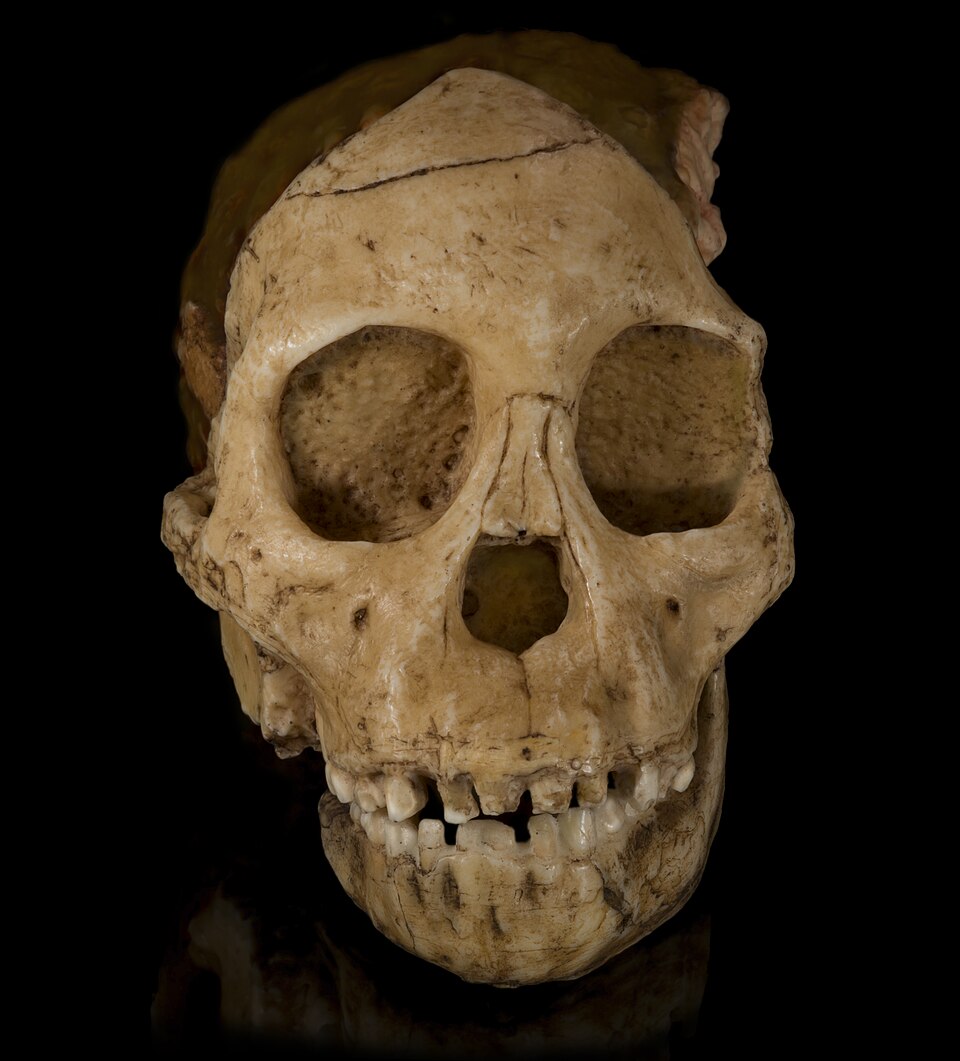
The Taung Child consists of three associated elements: a partial cranium with much of the face and right side of the braincase, a corresponding right hemi-mandible with deciduous dentition and erupting first molars, and a natural endocast preserving the impression of part of the cerebral surface. All three were recovered from the same block of pinkish calcrete matrix and clearly belong to a single juvenile individual.1, 10

_(Upper_Pliocene,_2.8_Ma;_quarry_near_Taung,_North_West_Province,_South_Africa).jpg){kind=link}
The face is small and orthognathic compared with extant great apes of similar age, with a shallow muzzle, a relatively flat nasal region, and modest canine sockets. The dental arcade is parabolic rather than U-shaped, and the deciduous canines do not project markedly above the occlusal plane.1, 14 The first permanent molars (M1) had only just erupted into occlusion at the time of death, with little or no occlusal wear, anchoring all subsequent attempts to estimate the child's age.3, 14
The basicranium preserves the foramen magnum in a position that is intermediate between that of extant great apes and that of modern humans, but distinctly closer to the human end of the range. Quantitative work on foramen magnum position across living mammals has confirmed that, in bipedal taxa, the foramen lies further forward and faces more directly downward than in habitually quadrupedal species, lending posthumous support to Dart's original locomotor inference.24 The natural endocast, finally, reproduces a portion of the right cerebral hemisphere—much of the temporal and parietal lobes and the lateral surface of the frontal lobe—along with venous sinus impressions that have been the focus of every subsequent reanalysis of the specimen's brain anatomy.10, 13
Cranial capacity and the lunate sulcus
Estimating the volume of the Taung Child's brain has occupied palaeoneurologists for a century. Dart's original 1925 estimate, derived from a wax cast of the natural endocast and adjusted for the missing left hemisphere, was approximately 520 cm³.1, 13 Later workers, applying different reconstruction methods and taking better account of the child's incomplete brain growth, produced lower figures. The most widely cited modern estimate is around 404 cm³, derived from independent reconstruction of the hemi-endocast and corroborated by digital reconstruction of the endocranial cavity from computed tomography data.10, 23 Across the published literature, estimates have ranged from approximately 382 cm³ to 530 cm³, depending on the assumed adult value and the technique used to fill in the missing portions.10, 22, 23

{kind=link}
Endocranial volume of the Taung Child: published estimates1, 10, 22, 23
The convergence of recent CT-based estimates around 400 cm³ places the Taung Child's brain in the same volumetric range as a modern adult chimpanzee. Because australopith brains had largely completed growth by the age at which Taung died, the small absolute size cannot be dismissed as a juvenile artefact: it is broadly representative of Australopithecus africanus adult endocranial volume, which falls in the range of approximately 400 to 560 cm³.10, 22
The more contentious question has always been the brain's organisation rather than its volume. Dart's claim that the lunate sulcus on the Taung endocast lay in a posterior, humanlike position implied an enlargement of the parietal association cortex at the expense of primary visual cortex—a hallmark of human neuroanatomy. The American palaeoneurologist Dean Falk challenged this identification in a series of papers beginning in the 1980s, arguing that the groove Dart had identified as the lunate sulcus was in fact a different structure and that Taung's true lunate sulcus, if visible at all, lay in an anterior, apelike position.12, 13 Ralph Holloway, working from his own latex casts, defended a more posterior position and rejected Falk's identification, and the resulting Falk–Holloway exchange ran across more than a decade of publications.12 Most contemporary palaeoneurologists conclude that no clear lunate sulcus can be unambiguously identified on the Taung endocast at all, and that arguments built on its supposed location must be treated with corresponding caution.10, 13
High-resolution CT imaging of the endocast
The most detailed examination of the Taung specimen came in 2012 and 2014, when high-resolution computed tomography of the partial cranium and natural endocast became feasible. In a 2012 PNAS paper, Falk and colleagues reported that the endocast preserved a small triangular remnant of the anterior fontanelle from which a metopic suture coursed forward along the midline. They argued that the late persistence of an open metopic suture in the Taung child paralleled a feature of modern human neonates and might reflect three coupled selective pressures in early hominins: an obstetric dilemma created by bipedalism, high postnatal brain growth rates, and reorganization of the frontal cortex.11
Two years later, Holloway, Broadfield, and Carlson published new high-resolution CT data of the same specimen in PNAS and reached a different conclusion. Their scans showed that, although a possible remnant of the metopic suture is present in the nasion–glabella region of the partial cranium, it does not extend along the frontal crest in the manner Falk and colleagues had described, and the supposed anterior fontanelle remnant is not visible in the CT data with the morphology required by their hypothesis.10 Holloway and colleagues concluded that, "if delayed metopic suture closure were adaptive...there is no evidence this mechanism evolved before Homo," and that Taung's brain growth pattern, as far as it can be reconstructed, was apelike rather than humanlike.10 Subsequent reanalyses of the endocast surface using larger comparative samples of living hominoids have generally supported a more conservative reading of the Taung neurology, in which the specimen documents brain expansion of only modest degree relative to its great-ape ancestry.10, 13
Age at death and dental development
Dart's original interpretation, based on the eruption pattern of the deciduous dentition and the just-erupting first molars, placed the Taung Child at approximately six years of age, applying a modern human dental schedule to the specimen.1 Subsequent work has progressively revised this estimate downward, as researchers have abandoned the human schedule in favour of methods anchored in the actual rates of enamel deposition recorded in the teeth themselves.
The first systematic CT-based assessment was performed by Glenn Conroy and Michael Vannier in 1987, using early medical CT scanners to image unerupted tooth crowns and roots inside the maxilla and mandible. They concluded that the dental development of the Taung skull was more rapid than that of modern humans and more consistent with an apelike pattern of growth.14 Two decades later, Lacruz, Ramirez Rozzi, and Bromage applied incremental enamel histology to the M1 of the Taung child, counting the perikymata (the surface manifestation of long-period growth lines) and combining the count with the growth rate of M1 crown formation observed in another A. africanus juvenile from Sterkfontein. They estimated the child's age at death at between 3.7 and 3.9 years, and they identified a band of linear enamel hypoplasia approximately 11 perikymata from the cervix on the first molar that recorded a brief episode of physiological stress recorded in the enamel during the final phase of crown formation—a possible illness, malnutrition event, or weaning crisis.15
Modern syntheses settle on an age of approximately 3 to 4 years at death, with most authorities favouring the lower end of that range.3, 15 The implication is that Australopithecus africanus dental development was faster than that of modern humans and more apelike in tempo—a pattern consistent with the species' small adult brain, short interbirth intervals expected in great apes, and absence of the prolonged childhood that characterises later Homo.
Eagle predation and the death of the child
For most of the twentieth century, the prevailing taphonomic story for the Taung assemblage held that the bones had been accumulated by carnivores—leopards, hyaenas, or extinct sabretooths—and that the Taung Child was a leopard kill carried back to a feeding site near a cave. Lee Berger and Ron Clarke offered a radically different interpretation in 1995 in the Journal of Human Evolution. Reanalysing the entire associated faunal assemblage from the Type Site, they noted that it was dominated by small-bodied animals—hyraxes, small bovids, juvenile primates, tortoises—rather than the large prey expected of mammalian carnivores, and that the bones bore patterns of damage strikingly similar to those left by modern large birds of prey on their prey items.6 They argued that a large raptor was the principal taphonomic agent for the Taung deposit and proposed that the Taung Child itself had been killed and partially consumed by such a bird.6
The hypothesis was initially controversial, in part because no obvious bird-of-prey damage had then been described on the Taung type cranium itself. Two developments in 2006 turned the tide. First, W. Scott McGraw and colleagues published a detailed taphonomic study of more than a thousand bones recovered from beneath sixteen nests of African crowned eagles (Stephanoaetus coronatus) in the Taï Forest of Côte d'Ivoire, documenting the precise patterns of damage—orbital punctures, raked breakage of long bones, V-shaped beak nicks—that crowned eagles inflict on their primate prey, including monkeys substantially larger than a four-year-old child.9 Second, in the same year and in the same journal, Berger published a re-examination of the Taung type-skull itself, identifying previously undescribed punctures and gouges in the orbital floors that closely matched the eagle damage documented by McGraw and colleagues.7
In 2007 Berger and McGraw together extended the case in the South African Journal of Science, presenting additional damage on the Taung cranium and showing that the orbital and basicranial gouges fall within the range of damage left by extant crowned eagles on monkey crania.8 Modern crowned eagles in African forests routinely take prey weighing six to ten kilograms, and small-bodied primates weighing more than twenty kilograms have been recorded among their kills. A juvenile australopith of three or four years would have fallen well within their capacity. The convergence of independent evidence—the small-bodied composition of the Taung faunal assemblage, the perforations and gouges on the type cranium, the close anatomical match to modern crowned-eagle damage—has made the eagle-predation hypothesis the standard taphonomic interpretation of the Taung type site.6, 7, 8, 9
Geology, depositional context, and dating
The Taung Child is unusual among South African australopith fossils in lacking a precisely known stratigraphic context. The original block of matrix containing the cranium was extracted from the Buxton–Norlim Limeworks during ordinary commercial quarrying, and the exact spot from which it came was destroyed before any geologist could record it. The site has therefore had to be reconstructed indirectly, by piecing together what is known of the surrounding tufa, calcrete, and breccia deposits at Buxton–Norlim.16, 17
For decades the Taung deposit was assumed, by analogy with Sterkfontein and other Cradle of Humankind sites, to be a brecciated cave fill formed by the collapse of dolomitic cave roofs. Reanalysis by Hopley and colleagues in 2013 challenged this paradigm. Using sedimentological and palaeomagnetic methods, they showed that the pink calcrete deposit from which the type specimen derives is a pedogenic carbonate—a calcrete soil—formed at or near the contemporary land surface rather than inside a cave.16 In this revised model, the Taung Child died not in a buried cavern but in an open landscape of tufa-fed wetlands and calcareous soils, the kind of paleosol setting more characteristic of East African hominin localities than of the dolomitic cave deposits of the Cradle of Humankind further east.16, 17
The age of the deposit is also imprecise. Estimates have ranged across more than a million years of late Pliocene and earliest Pleistocene time. The Smithsonian Human Origins Program lists the Taung specimen at approximately 2.8 million years old, while Britannica gives a range of 2.3 to 2.8 million years, both based on biochronological correlation of the associated mammalian fauna.3, 5 A 2025 analysis by J. F. Thackeray, applying biochronological dimensions of the lower first molar, places the specimen at approximately 2.58 million years—a figure that essentially coincides with the formal Plio–Pleistocene boundary defined by the Gauss–Matuyama palaeomagnetic reversal.18 The renewed investigations at Buxton–Norlim led by Brian Kuhn, Andy Herries, and colleagues since 2010 are continuing the geochronological work, applying combined uranium–lead and palaeomagnetic methods to constrain the depositional history of the broader limeworks.17
Key dates and figures associated with the Taung Child1, 2, 3, 10, 15, 18
| Item | Value | Source |
|---|---|---|
| Discovery (face freed from matrix) | 23 December 1924 | Dart 1925; Smithsonian HOP |
| Publication of Nature paper | 7 February 1925 | Dart 1925 |
| Type locality | Buxton–Norlim Limeworks, Taung, North West Province, South Africa | Dart 1925; Hopley et al. 2013 |
| Age at death | ~3.7–4 years | Lacruz et al. 2005; Smithsonian HOP |
| Modern endocranial volume estimate | ~382–404 cm³ | Holloway 2007; Holloway et al. 2014 |
| Dart's 1925 endocranial volume estimate | ~520 cm³ | Dart 1925 |
| Geological age (biochronology) | ~2.58–2.8 Ma | Smithsonian HOP; Thackeray 2025 |
| Probable cause of death | Predation by a large raptor (likely Stephanoaetus coronatus) | Berger & Clarke 1995; Berger 2006; Berger & McGraw 2007 |
| Current repository | University of the Witwatersrand, Johannesburg | Wits University Evolutionary Studies Institute |
Legacy and continuing significance
The Taung Child remains the type specimen of Australopithecus africanus a hundred years after Dart's announcement, and it remains one of the most studied early hominin fossils ever recovered.2, 26 Several broad points are now widely accepted that were not in 1925. First, Dart was substantially correct about the specimen's intermediate position: the dental and basicranial features that he identified as hominin are confirmed by every reanalysis since, and his locomotor inference from the foramen magnum has been validated by quantitative comparative work on living mammals.2, 24 Second, his more ambitious neurological claims have not survived as well: the Taung endocast has not yielded unambiguous evidence of either a posteriorly placed lunate sulcus or a delayed metopic closure of human-like form, and current reconstructions place the absolute brain volume firmly within the great-ape range.10, 13
Third, the discovery of Sahelanthropus tchadensis, Orrorin tugenensis, and the genus Ardipithecus—all dating to between roughly 4.4 and 7 million years ago—has pushed the hominin fossil record more than four million years deeper than the Taung Child, leaving Australopithecus africanus as a well-documented but no longer pioneering inhabitant of the human family tree.2, 4, 25 The species itself is now known from rich samples at Sterkfontein, Makapansgat, and Gladysvale, with adult cranial capacities ranging from approximately 400 to 560 cm³ and a growing body of postcranial material documenting habitual bipedalism alongside retained climbing adaptations.4, 22, 25
What endures from 1925 is not the specific neurological argument but the structural insight: that a small-brained, small-toothed, bipedal ape could be the ancestor of modern humans, and that the search for that ancestor belonged in Africa. The Taung Child shifted the geographical centre of palaeoanthropology from Europe and Asia to the African continent, broke the model of brain-first human evolution, and established the genus that, in different forms, still anchors the deepest reaches of the human family tree.2, 25, 26 The fossil itself now sits in the Fossil Primate and Hominid Vault of the University of the Witwatersrand's Evolutionary Studies Institute in Johannesburg, where it was first prepared, and where the renewed investigations at Buxton–Norlim and the continuing reanalyses of its endocast and dentition ensure that, a century on, it has not yet finished speaking.17, 26
References
Dart and the Taung juvenile: making sense of a century-old record of hominin evolution in Africa
Brief communication: Predatory bird damage to the Taung type-skull of Australopithecus africanus Dart 1925
Primate remains from African crowned eagle (Stephanoaetus coronatus) nests in Ivory Coast's Tai Forest: implications for primate predation and early hominid taphonomy in South Africa
New high-resolution computed tomography data of the Taung partial cranium and endocast and their bearing on metopism and hominin brain evolution
Metopic suture of Taung (Australopithecus africanus) and its implications for hominin brain evolution
The Taung endocast and the lunate sulcus: a rejection of the hypothesis of its anterior position
The natural endocast of Taung (Australopithecus africanus): insights from the unpublished papers of Raymond Arthur Dart
Brief communication: beyond the South African cave paradigm—Australopithecus africanus from Plio-Pleistocene paleosol deposits at Taung
Renewed investigations at Taung; 90 years after the discovery of Australopithecus africanus
The Taung Child (Australopithecus africanus), the Plio-Pleistocene boundary and a supernova hypothesis
Endocranial volume of Australopithecus africanus: new CT-based estimates and the effects of missing data and small sample size
Contesting a legendary legacy: a century of reflection on Raymond Dart and the Taung skull