Overview
- Anomalocaris was the largest known predator of the Cambrian seas, reaching body lengths of up to one meter, and belongs to the Radiodonta, a group of stem-lineage arthropods characterized by a pair of large frontal appendages, a circular mouth apparatus, and laterally arranged swimming flaps.
- First discovered in the 1890s as isolated body parts mistaken for three separate organisms — a shrimp (the frontal appendage), a jellyfish (the circular mouth), and a sponge (the body) — Anomalocaris was not recognized as a single animal until Harry Whittington and Derek Briggs reassembled it in 1985.
- Exceptionally preserved compound eyes containing over 16,000 lenses per eye, discovered in Cambrian deposits of the Emu Bay Shale in Australia, demonstrate that Anomalocaris possessed among the most sophisticated visual systems of any Cambrian animal, comparable in acuity to modern dragonflies.
Anomalocaris ("anomalous shrimp") is an extinct genus of large marine animal from the Cambrian period, approximately 520 to 505 million years ago, and the most famous member of the Radiodonta, a group of stem-lineage arthropods that were among the earliest large predators in the history of animal life.1, 10 The genus is known primarily from exceptionally preserved fossils in the Burgess Shale of British Columbia, Canada (approximately 508 million years old), and the Chengjiang biota of Yunnan Province, China (approximately 518 million years old), two of the most important Cambrian fossil deposits in the world.1, 2, 11 At up to one meter in body length, Anomalocaris dwarfed nearly all other Cambrian animals and likely occupied the apex predator niche in Cambrian marine ecosystems, making it a key organism for understanding the origins of complex food webs and predator-prey dynamics in early animal evolution.1, 7
{kind=link}
Discovery and reassembly
The history of Anomalocaris is one of the most celebrated tales of misidentification in paleontology. The first fossils attributable to the genus were discovered in the late nineteenth century by Joseph Frederick Whiteaves, who in 1892 described an isolated frontal appendage from the Cambrian of British Columbia as the abdomen of a crustacean, naming it Anomalocaris canadensis, the "anomalous shrimp of Canada," because it lacked the features expected of a typical crustacean tail.1, 4 The circular mouth apparatus, when found separately, was described as a jellyfish and given the name Peytoia. The flattened body, preserved without appendages, was interpreted as a sponge and named Laggania.1, 4 For nearly a century, what was actually a single spectacular animal was scattered across the taxonomic literature as three unrelated organisms.4
The true nature of Anomalocaris was revealed in 1985, when Harry Whittington and Derek Briggs published a landmark reassessment based on complete or near-complete specimens from the Burgess Shale.1 Working with specimens that preserved the frontal appendages, circular mouth, and body together in articulation, Whittington and Briggs demonstrated that the "shrimp," "jellyfish," and "sponge" were all parts of a single, very large animal unlike anything in the modern oceans.1 The reassembled Anomalocaris was immediately recognized as the largest known animal of the Cambrian, and its description electrified the paleontological community, providing a vivid example of how incomplete preservation can lead to fundamental misunderstanding of extinct organisms.1, 4 Stephen Jay Gould made the reassembly of Anomalocaris a centerpiece of his 1989 book Wonderful Life, which brought the Burgess Shale fauna to popular attention and used Anomalocaris to illustrate the radical strangeness of Cambrian life.4
Body plan and morphology
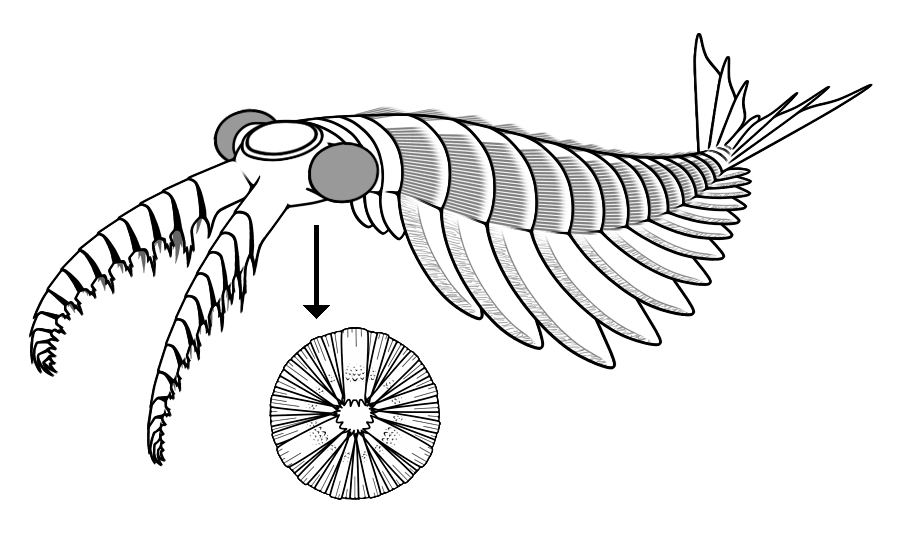
The body plan of Anomalocaris is distinctive and has no close analogue among living animals. The body was elongated, laterally compressed, and bore a series of paired, overlapping lateral lobes or flaps along each side, which are interpreted as swimming structures that propelled the animal through the water column by means of undulating, wave-like motions, similar in principle to the movement of modern cuttlefish or rays.1, 16 The head bore a pair of large, segmented frontal appendages, each composed of multiple articulating podomeres armed with spines along their inner margins, that were positioned in front of and below the mouth.1, 16 These appendages were flexible and capable of curling inward, and they are widely interpreted as grasping organs used to capture and manipulate prey before delivering it to the mouth.1, 7
The mouth was located on the ventral surface of the head and consisted of a circular or sub-circular oral cone composed of 32 overlapping plates arranged in a ring, with larger plates alternating with smaller ones in a pattern reminiscent of a pineapple slice.1, 14 The plates converged toward a central opening but did not completely close, leaving a small gap at the center.14 The question of whether this mouth apparatus could generate enough force to crack trilobite exoskeletons has been debated. Allison Daley and Graham Budd noted that the oral cone could not fully close and likely could not crush hard shells, suggesting that Anomalocaris may have focused on softer prey or on damaged and molting trilobites rather than attacking intact, heavily armored individuals.7
The tail region bore a fan-shaped caudal structure composed of radiating fin-like elements, which likely served as a stabilizer and rudder during swimming.1, 16 The overall body plan is that of an active, nektonic predator capable of sustained swimming, a sharp contrast to the predominantly benthic and slow-moving fauna that characterized most Cambrian marine communities.1, 7
Compound eyes and visual acuity
Among the most striking features of Anomalocaris are its large, stalked compound eyes. In 2011, John Paterson and colleagues described a pair of exceptionally preserved isolated compound eyes from the Early Cambrian Emu Bay Shale of Kangaroo Island, South Australia, attributed to Anomalocaris on the basis of their size, association, and morphology.3 Each eye measured approximately 3 centimeters in length and contained over 16,000 individual hexagonally packed lenses, a number that exceeds the lens count of most living arthropods and approaches that of modern dragonflies, which are among the most visually acute insects alive today.3
The high lens count and tightly packed arrangement of the ommatidial units indicate that Anomalocaris possessed excellent visual acuity, sufficient to detect and track moving prey from a considerable distance in the water column.3 Paterson and colleagues argued that the presence of such sophisticated compound eyes in an Early Cambrian predator supports the "arms race" hypothesis for the Cambrian explosion, which posits that the evolution of vision-driven predation was a key trigger for the rapid diversification of animal body plans, as prey lineages evolved increasingly elaborate defenses (shells, spines, burrowing behavior) in response to the new threat of visual predators.3
The eyes of Anomalocaris also bear on the broader question of arthropod evolution. Compound eyes of this type, composed of individual optical units (ommatidia) packed into a convex array, are a defining feature of living arthropods, from insects to crustaceans to horseshoe crabs. The discovery of compound eyes with modern-type optics in a stem-lineage arthropod from the Early Cambrian demonstrates that the compound eye had already evolved to a high level of sophistication before the divergence of the major arthropod groups, placing its origin deep in the Cambrian or possibly even earlier.3
Ecological role and feeding strategies
The traditional interpretation of Anomalocaris as a trilobite predator is based on several lines of evidence. Healed bite marks on trilobite exoskeletons from Cambrian deposits have been attributed to Anomalocaris attack, and the size and shape of some of these injuries are broadly consistent with the dimensions of anomalocaridid frontal appendages and oral cones.13 Christopher Nedin described W-shaped bite marks on trilobites from the Emu Bay Shale and argued that they were produced by the oral cone of Anomalocaris.13 However, the interpretation has been questioned, with Daley and Budd pointing out that the inability of the oral cone to fully close makes it unlikely that Anomalocaris could have bitten cleanly through mineralized trilobite exoskeletons, and suggesting instead that the animals may have processed prey by flexing and cracking it with their frontal appendages before ingesting the soft internal tissues.7
The discovery of suspension-feeding radiodonts has expanded understanding of the ecological diversity within this group. In 2014, Jakob Vinther and colleagues described Tamisiocaris borealis from the Early Cambrian Sirius Passet deposit of Greenland, a radiodont whose frontal appendages bore fine, densely spaced setae rather than robust spines, interpreted as a filtering apparatus for capturing small planktonic organisms.6 This demonstrated that radiodonts were not exclusively predatory but had diversified into multiple trophic niches by the Early Cambrian, paralleling the ecological range seen in modern marine arthropod groups such as decapod crustaceans.6 The genus Hurdia, another Burgess Shale radiodont described in detail by Daley and colleagues, possessed a large frontal carapace element and appendages with intermediate spine morphology, suggesting yet another distinct feeding strategy within the group.5, 8
As the largest nektonic predator in Cambrian oceans, Anomalocaris almost certainly exerted strong top-down selective pressure on its prey communities, shaping the evolution of defensive adaptations in trilobites, brachiopods, and other benthic organisms.7 The evolutionary arms race between anomalocaridid predators and their prey is thought to have been one of the driving forces behind the rapid elaboration of shells, spines, and other defensive structures that characterize the Cambrian fossil record.3, 7
Radiodont diversity and phylogeny
The order Radiodonta, to which Anomalocaris belongs, is now recognized as a diverse and widespread group of stem-lineage arthropods that persisted from the Early Cambrian into at least the Ordovician period, a temporal range of over 80 million years.9, 10 In addition to Anomalocaris, the group includes genera such as Hurdia, Peytoia (now recognized as distinct from the oral cone originally assigned to Anomalocaris), Amplectobelua, Tamisiocaris, and Aegirocassis, among many others, spanning a wide range of body sizes and ecological strategies.5, 6, 8, 9
The largest known radiodont is Aegirocassis benmoulai from the Early Ordovician of Morocco, described by Peter Van Roy and Derek Briggs in 2011, which reached an estimated body length of approximately two meters, making it one of the largest animals of the entire early Paleozoic.9 Like Tamisiocaris, Aegirocassis was a suspension feeder, demonstrating that the filter-feeding body plan within Radiodonta was both successful and long-lived.9 The persistence of radiodonts into the Ordovician and their ecological diversification into multiple trophic roles indicate that the group was far more successful and enduring than previously appreciated, not merely a short-lived Cambrian experiment but a significant component of early Paleozoic marine ecosystems.9, 10
The phylogenetic position of Radiodonta is critical for understanding arthropod evolution. Cladistic analyses consistently place radiodonts as stem-group euarthropods, meaning they branched off before the evolution of the fully arthrodized, segmented body plan with jointed legs that characterizes crown-group arthropods (insects, crustaceans, arachnids, myriapods).5, 10, 15 Recent work by Joseph Moysiuk and Jean-Bernard Caron has refined this picture by identifying new Burgess Shale radiodonts that preserve features transitional between the radiodont body plan and that of more derived arthropods, including structures that may be homologous to the mandibles of modern crustaceans and insects.15 These findings position Anomalocaris and its relatives not as evolutionary oddities but as key waypoints in the assembly of the arthropod body plan, the most successful animal body plan in the history of life on Earth.10, 15
Significance for Cambrian paleobiology
Anomalocaris has become an icon of the Cambrian explosion, the geologically rapid appearance of most major animal body plans in the fossil record beginning approximately 538.8 million years ago. Its significance extends beyond its status as the largest Cambrian predator. The tortuous history of its discovery and reconstruction illustrates the profound difficulty of interpreting organisms with no modern analogues from incomplete fossil material, a challenge that pervades the study of Cambrian life.1, 4 The fact that an animal as large and distinctive as Anomalocaris could remain unrecognized for nearly a century despite being represented by hundreds of specimens in museum collections is a sobering reminder of how much of the Cambrian fauna may remain misunderstood or unrecognized in existing collections.4
The ecological implications of Anomalocaris are equally profound. Its presence at the top of Cambrian food webs demonstrates that complex trophic structures, including multi-level food chains with apex predators, were established very early in the history of animal life.7 The sophistication of its visual system, its active swimming capabilities, and the diversity of feeding strategies within Radiodonta all point to a Cambrian ocean that was far more ecologically complex than the simple, predator-free world once imagined for the earliest animal communities.3, 6, 7 Together with the broader Burgess Shale and Chengjiang faunas of which it is a part, Anomalocaris provides critical evidence for understanding how the modern marine biosphere, with its complex ecosystems and interlocking trophic relationships, first came into being.4, 7
References
Anomalocaris and other large animals in the Lower Cambrian Chengjiang fauna of southwest China
Acute vision in the giant Cambrian predator Anomalocaris and the origin of compound eyes
The Burgess Shale anomalocaridid Hurdia and its significance for early euarthropod evolution
Morphology and systematics of the anomalocaridid arthropod Hurdia from the Middle Cambrian of British Columbia
Amplectobelua symbrachiata and the earliest evidence of suspension feeding in stem-group euarthropods
Were early trilobites stressed by Anomalocaris? Isotopic and microstructural evidence
An exceptionally preserved three-dimensionally articulated oral cone of Anomalocaris from the Cambrian Emu Bay Shale
A new radiodontan from the Burgess Shale reveals an early Cambrian origin of the mandibulate appendage