Overview
- Mammals during the Mesozoic era were far more ecologically diverse than the traditional “small, nocturnal insectivore” stereotype suggests, with fossil discoveries revealing species adapted for gliding, swimming, burrowing, climbing, and even feeding on small dinosaurs.
- Key discoveries including Castorocauda (a swimming, beaver-like mammal from the Middle Jurassic), Volaticotherium (a gliding mammal from the Late Jurassic or Early Cretaceous), and Repenomamus (a badger-sized predator that ate juvenile dinosaurs) have fundamentally revised understanding of Mesozoic mammalian ecology.
- Molecular and morphological phylogenetic analyses suggest that the crown-group divergence of modern mammalian orders occurred during the Cretaceous, well before the end-Cretaceous extinction, meaning that the ecological foundations for the mammalian radiation were laid while dinosaurs still dominated terrestrial ecosystems.
For much of the twentieth century, the conventional picture of Mesozoic mammals was one of ecological marginality: small, nocturnal, insectivorous creatures living in the shadow of the dinosaurs, constrained to a narrow range of body sizes and ecological roles until the end-Cretaceous extinction liberated them to diversify.5, 8 This view, while not entirely without basis, has been comprehensively overturned by a wave of fossil discoveries since the late 1990s, predominantly from Mesozoic deposits in China, Mongolia, and South America, that reveal a Mesozoic mammalian fauna of surprising ecological breadth and morphological innovation.5, 16 Mammals during the age of dinosaurs included gliders, swimmers, burrowers, climbers, and even predators large enough to eat juvenile dinosaurs, occupying ecological niches previously thought to have been available only after the disappearance of non-avian dinosaurs.1, 2, 3
{kind=link}
Aquatic and gliding forms
The discovery of Castorocauda lutrasimilis from the Middle Jurassic Daohugou Biota of Inner Mongolia (approximately 164 million years ago) was a landmark in the reassessment of Mesozoic mammalian ecology. Described by Qiang Ji and colleagues in 2006, Castorocauda was a docodont mammaliaform approximately 42 centimeters in total length, with a broad, flattened, beaver-like tail, webbed hind feet, and dense fur preserved as carbonized impressions in the fine-grained lake sediments.1 The morphology of its limbs and tail strongly suggests a semi-aquatic lifestyle comparable to that of modern platypuses or otters, indicating that mammals had invaded freshwater habitats by the Middle Jurassic, more than 100 million years earlier than previously documented.1, 5
Equally unexpected was the discovery of Volaticotherium antiquum, a gliding mammal from Mesozoic deposits of Inner Mongolia described by Jin Meng and colleagues in 2006.2 Volaticotherium possessed a furred gliding membrane (patagium) extending between its forelimbs and hind limbs, preserved as a carbon film in the slab, making it the earliest known gliding mammal. The animal weighed an estimated 70 grams and bore elongated limbs with specialized proportions consistent with aerial locomotion, demonstrating that mammals had independently evolved gliding flight during the Mesozoic, paralleling later independent origins of gliding in flying squirrels, colugos, and sugar gliders.2 Subsequent discoveries from the Jurassic of China, including additional gliding mammaliaforms such as Maiopatagium and Vilevolodon, confirmed that gliding evolved multiple times among Mesozoic mammalian lineages.4
Predators and burrowers
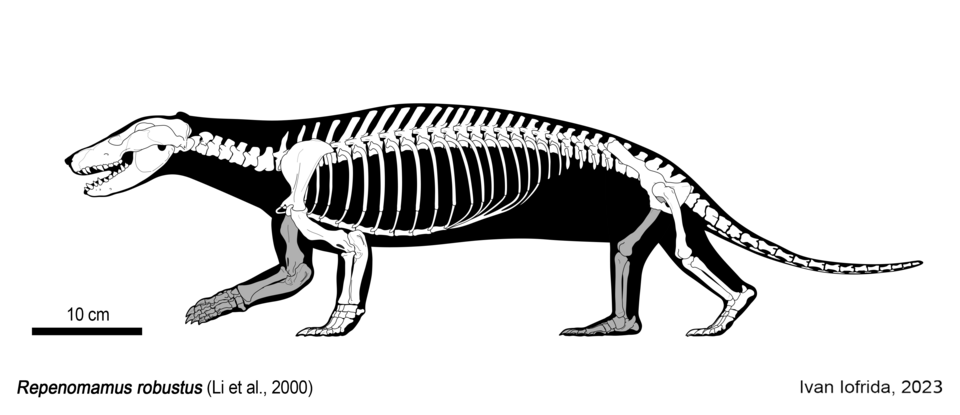
Perhaps the most dramatic challenge to the "small insectivore" paradigm came with the description of Repenomamus giganticus and Repenomamus robustus from the Early Cretaceous Yixian Formation of Liaoning Province, China. Yaoming Hu and colleagues reported in 2005 that R. giganticus was a triconodont mammal with an estimated body mass of 12 to 14 kilograms, making it the largest known Mesozoic mammal, roughly the size of a modern badger.3 More remarkably, a specimen of the smaller R. robustus (approximately 4 to 6 kilograms) preserved the partially digested skeleton of a juvenile Psittacosaurus, a small ceratopsian dinosaur, within its abdominal cavity, providing direct evidence that at least some Mesozoic mammals preyed on dinosaurs.3 The robust skull, powerful jaw musculature, and non-specialized dentition of Repenomamus indicated a generalized carnivorous or omnivorous diet, and its body size exceeded that of many contemporaneous dinosaur species.3, 8
Fossorial (burrowing) adaptations have also been documented in Mesozoic mammals. Fruitafossor windscheffeli from the Late Jurassic Morrison Formation of Colorado possessed robust, shovel-like forelimbs and simplified, peg-like teeth resembling those of modern armadillos, interpreted as adaptations for digging and feeding on colonial insects.5 The Early Cretaceous Spinolestes xenarthrosus from the Las Hoyas deposit of Spain preserved remarkable soft tissue including fur, spines similar to modern hedgehog quills, and a diaphragm, providing evidence of both burrowing behavior and advanced thermoregulatory adaptations.13 These fossorial forms demonstrate that Mesozoic mammals occupied subterranean niches that placed them outside the direct competitive arena with most dinosaurs.5, 13
Morphological disparity and body size
Quantitative analyses of morphological disparity, measured by the range and variance of skeletal proportions across species, have confirmed that Mesozoic mammals were far more ecomorphologically diverse than body size alone would suggest. Zhe-Xi Luo's comprehensive 2007 review demonstrated that Mesozoic mammaliaforms occupied a morphospace comparable in breadth to that of modern small mammals, with diversity in dental morphology, limb proportions, and locomotor adaptations that rivaled the range seen among living insectivores, rodents, and small carnivores.5 Dental evolution was particularly innovative: eutriconodonts evolved interlocking shearing teeth for carnivory, multituberculates developed complex, multirowed teeth for herbivory and granivory that functioned analogously to rodent incisors and premolars, and docodonts evolved diverse molar patterns suited for omnivory and insectivory.5, 8
Body size, however, was genuinely constrained during the Mesozoic relative to the Cenozoic. With the exception of Repenomamus and a few other outliers, most Mesozoic mammals weighed between 2 grams and 1 kilogram, and the largest known species were roughly two orders of magnitude smaller than the largest Cenozoic mammals.5, 8 This size constraint has been attributed to competitive exclusion by small and medium-sized dinosaurs occupying the same body-size range, an interpretation supported by the observation that the mammalian body-size range expanded rapidly after the end-Cretaceous extinction removed dinosaurian competitors.5, 10 The ecological diversity of Mesozoic mammals was thus achieved primarily through differentiation of locomotor mode, diet, and habitat rather than through the body-size disparity that characterizes the Cenozoic mammalian radiation.5, 14
Phylogenetic implications
The discovery of ecologically diverse Mesozoic mammals has been accompanied by major advances in understanding mammalian phylogeny. Molecular clock estimates and increasingly complete fossil records suggest that the divergence of the three living mammalian subclasses, Monotremata (egg-laying mammals), Metatheria (marsupials and their relatives), and Eutheria (placentals and their relatives), occurred during the Jurassic or Early Cretaceous, much earlier than the oldest definitive crown-group fossils would suggest on their own.7, 9 The discovery of Sinodelphys szalayi from the Early Cretaceous Yixian Formation, described as the oldest known metatherian, and Eomaia scansoria from the same deposits, described as one of the oldest known eutherians, pushed the documented fossil record of the two major living lineages back to approximately 125 million years ago.9, 15
These early members of the eutherian and metatherian lineages were themselves small and relatively generalized, but their presence in the Early Cretaceous implies that much of the phylogenetic framework for the later mammalian adaptive radiation was already in place while dinosaurs still dominated terrestrial ecosystems.9, 10 The evolution of the middle ear, a hallmark of mammalian anatomy involving the detachment of the post-dentary bones from the jaw and their incorporation into the auditory apparatus, appears to have occurred independently in multiple Mesozoic lineages, as demonstrated by Luo and colleagues' analysis of Cretaceous fossils preserving transitional ear morphologies.12 This convergent evolution of a key mammalian feature underscores the remarkable evolutionary dynamism of Mesozoic mammals, which were experimenting with and independently arriving at innovations that define the modern mammalian body plan.5, 12
Reassessing the Mesozoic
The cumulative effect of these discoveries has been to fundamentally revise the narrative of mammalian evolution. Rather than a prolonged period of evolutionary stasis punctuated by an explosive radiation only after the disappearance of the dinosaurs, the Mesozoic mammalian record now reveals a dynamic evolutionary history in which mammals continuously explored new ecological opportunities, evolved novel body plans, and diversified into a range of habitats and feeding strategies despite the presence of dinosaurian competitors.5, 16 The post-Cretaceous radiation was not a creation from nothing but an expansion of possibilities that had been developing over more than 150 million years of Mesozoic evolution. What the end-Cretaceous extinction provided was not ecological novelty but ecological space, the removal of competitors that had constrained mammalian body size and limited access to diurnal, large-bodied, and megaherbivore niches.5, 8, 10 Understanding this Mesozoic prelude is essential for explaining why the mammalian radiation proceeded as rapidly and diversely as it did when the opportunity finally arose.8, 16
References
A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals
A new docodontan mammaliaform from the Late Jurassic of the Junggar Basin in Northwest China
New Jurassic mammaliaform sheds light on early evolution of mammal-like body proportions
Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary