Overview
- Spinosauridae is a family of large theropod dinosaurs within Megalosauroidea distinguished by elongated crocodile-like snouts, conical teeth, enlarged manual claws, and tall dorsal neural spines, with the group divided into the subfamilies Baryonychinae and Spinosaurinae.
- Multiple independent lines of evidence, including oxygen isotope ratios, bone microstructure showing unusually dense compact bone, and the discovery of a paddle-shaped tail in Spinosaurus, support semi-aquatic habits and piscivorous feeding ecology for at least some spinosaurids.
- The history of Spinosaurus research is uniquely dramatic: Ernst Stromer's original Egyptian fossils were destroyed in the 1944 Allied bombing of Munich, and the animal was not substantially known again until new North African material was described beginning in 2014, fundamentally transforming understanding of this group.
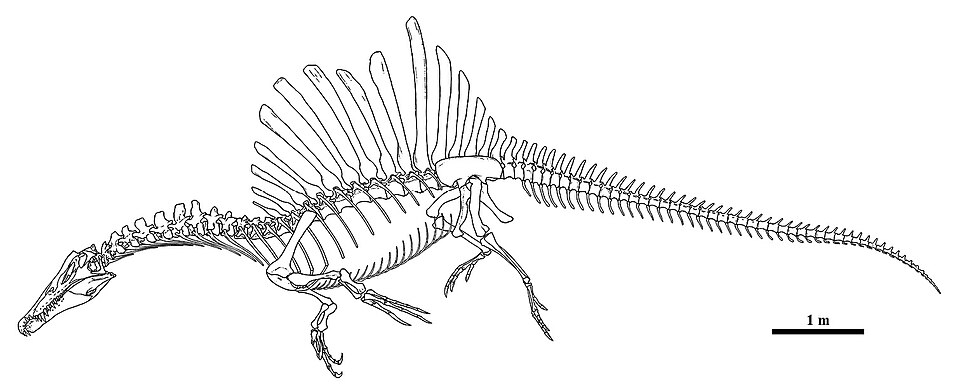
Spinosauridae is a family of large-bodied theropod dinosaurs characterised by elongated, narrow snouts bearing conical teeth, enlarged thumb claws, and tall neural spines on the dorsal vertebrae that formed a prominent sail or fin along the back. Spanning the Early to mid-Cretaceous period (approximately 130 to 95 million years ago), spinosaurids occupied a unique ecological niche among predatory dinosaurs, with multiple lines of evidence indicating that at least some members of the family were semi-aquatic and fed primarily on fish — a lifestyle without close parallel among other non-avian dinosaurs.7, 8 The group includes some of the largest theropods ever to have lived: Spinosaurus aegyptiacus of North Africa is estimated to have reached body lengths of 14 to 15 metres, rivalling or exceeding Tyrannosaurus rex in overall size, though with a markedly different body plan optimised for life at the water's edge rather than on open land.7
{kind=link}
The scientific history of Spinosauridae is as remarkable as the animals themselves. The type genus, Spinosaurus, was first described from Egyptian fossils in 1915, only for the original material to be destroyed during the Allied bombing of Munich in 1944. For decades the group remained poorly understood, until a cascade of discoveries beginning in the 1980s — most notably the English Baryonyx, the Nigerien Suchomimus, and new North African Spinosaurus material — transformed spinosaurids from an obscure curiosity into one of the most intensively studied dinosaur families in modern palaeontology.2, 4, 6
Discovery and historical overview
The story of Spinosauridae begins in the Western Desert of Egypt. In 1912, the German fossil collector Richard Markgraf excavated a partial skeleton from Cenomanian-aged (approximately 97 million years old) strata of the Bahariya Formation at Gebel el-Dist, near the Bahariya Oasis. The material was shipped to Munich, where the German palaeontologist Ernst Stromer von Reichenbach studied and described it in 1915, erecting the new genus and species Spinosaurus aegyptiacus — the "Egyptian spine lizard" — on the basis of an incomplete dentary, several vertebrae with enormously elongated neural spines reaching nearly two metres in height, and fragmentary limb elements.1 Stromer recognised that Spinosaurus was unlike any theropod previously known, noting the conical, unserrated teeth and the extraordinary dorsal sail, but the fragmentary nature of the holotype left many anatomical questions unanswered.
Those questions became permanently unresolvable on the night of 24–25 April 1944, when a Royal Air Force bombing raid struck the Bayerische Staatssammlung für Paläontologie und Geologie in Munich, destroying Stromer's irreplaceable holotype specimen (BSP 1912 VIII 19) along with other North African dinosaur material he had described, including the holotypes of the sauropod Aegyptosaurus and the theropod Carcharodontosaurus.6 Stromer had attempted to have the specimens moved to safety earlier in the war but was overruled by the museum's director. For decades afterward, knowledge of Spinosaurus rested primarily on Stromer's published descriptions and illustrations. It was not until 2006 that rediscovered photographs from Stromer's personal archive, donated by his son to the Munich collection in 1995, provided new anatomical information about the destroyed holotype, including previously unpublished views of the mandible and the mounted specimen.6
The modern understanding of spinosaurid anatomy began with the discovery of Baryonyx walkeri in 1983 by amateur fossil collector William Walker, who recovered an enormous manual claw from a clay pit in the Wealden Group of Surrey, England. Charig and Milner's preliminary description in Nature in 1986, followed by their comprehensive 1997 monograph, revealed a large theropod (approximately 7.5 to 10 metres long) with a strikingly elongated, narrow skull bearing numerous small conical teeth, a pronounced subrostral notch between the premaxilla and maxilla reminiscent of the rosette at the tip of a gharial's snout, and the largest known theropod manual claw — features that immediately suggested a piscivorous feeding strategy. Crucially, the abdominal region of the holotype preserved acid-etched scales of the fish Scheenstia mantelli and partially digested bones of a juvenile iguanodontid, providing the first direct evidence of diet in any spinosaurid.2, 3
In 1998, Paul Sereno and colleagues described Suchomimus tenerensis from the Aptian-aged (approximately 112 million years old) Elrhaz Formation of the Ténéré Desert, Niger. Known from a substantially complete skeleton approximately 11 metres long, Suchomimus confirmed the spinosaurid body plan on a larger scale: an elongated snout with a terminal rosette, conical teeth, a hypertrophied thumb claw, and moderately tall neural spines forming a low dorsal ridge.4 The description of Suchomimus established the broad framework of spinosaurid phylogeny, placing it within a Gondwanan radiation of fish-eating megalosauroids and demonstrating that the group was far more diverse and widespread than the fragmentary record of Spinosaurus alone had suggested.
Phylogenetic relationships
Spinosauridae is nested within Megalosauroidea, a basal tetanuran clade that also includes the Megalosauridae (containing Megalosaurus, Torvosaurus, and their relatives). The most comprehensive phylogenetic analysis of basal tetanuran relationships, published by Carrano, Benson, and Sampson in 2012 using 353 morphological characters across 61 taxa, confirmed the monophyly of Megalosauroidea and recovered Spinosauridae and Megalosauridae as sister groups, with the more basal Piatnitzkysauridae forming the outgroup to both.5 Megalosauroidea itself is the sister clade to all remaining tetanurans, including Allosauroidea and Coelurosauria, placing spinosaurids relatively basally within the broader theropod family tree — far removed from the coelurosaurian lineage that includes tyrannosaurids and birds.
Within Spinosauridae, phylogenetic analyses consistently recover two subfamilies: Baryonychinae and Spinosaurinae. Baryonychinae includes Baryonyx walkeri from the Barremian of England, Suchomimus tenerensis from the Aptian of Niger, and the recently described Isle of Wight taxa Ceratosuchops inferodios and Riparovenator milnerae. Baryonychines are generally characterised by a relatively greater number of teeth (including finely serrated premaxillary teeth in some taxa), a somewhat less extreme snout elongation compared to spinosaurines, and moderately tall neural spines.3, 4, 13 Spinosaurinae includes Spinosaurus aegyptiacus of North Africa and the South American taxa Irritator challengeri and Oxalaia quilombensis. Spinosaurines tend to have more extreme cranial elongation, fewer and entirely unserrated teeth, and the tallest neural spines in the family.15
A large-scale phylogenetic analysis by Barker and colleagues in 2021, incorporating the two new Isle of Wight spinosaurids and employing both parsimony and Bayesian methods, recovered a topology suggesting that Spinosauridae may have originated in Europe during the Early Cretaceous, with at least two independent dispersal events into Africa giving rise to the African baryonychines (such as Suchomimus) and the spinosaurines (such as Spinosaurus).13 This hypothesis of a European origin challenged earlier assumptions that the group was primarily Gondwanan, though the geographic sampling of spinosaurid fossils remains incomplete and the biogeographic picture continues to evolve with new discoveries.
Sales and Schultz's 2017 analysis of Brazilian spinosaurid cranial material further complicated the subfamilial picture by finding that some Brazilian taxa exhibit intermediate character states between Baryonychinae and Spinosaurinae, raising the possibility that Baryonychinae as traditionally defined may not be monophyletic.15 The internal phylogeny of Spinosauridae thus remains an active area of research, with each new discovery refining understanding of relationships within the family.
Distinctive morphology
Spinosaurids are among the most morphologically distinctive of all theropod dinosaurs, departing from the typical large-theropod body plan in several striking ways. The skull is the most immediately diagnostic element. Unlike the deep, laterally compressed skulls of most large theropods, spinosaurid skulls are elongated and dorsoventrally narrow, with a tubular snout that tapers anteriorly. The premaxillae bear a characteristic terminal rosette — a laterally expanded cluster of teeth at the tip of the snout, separated from the maxillary tooth row by a pronounced notch or constriction — that closely parallels the snout morphology of modern gharials (Gavialis gangeticus) and other longirostrine crocodilians.3, 12 The teeth themselves are conical and subcircular in cross-section, lacking the ziphodont (blade-like, serrated) morphology typical of most carnivorous theropods, and instead resembling the teeth of fish-eating crocodilians and certain predatory marine reptiles.4, 15
Finite element analysis and beam-theory modelling of spinosaurid rostra have demonstrated that the elongated snout was well suited to resisting the dorsoventrally directed bending forces generated during the rapid lateral striking motions used to capture fish, but was relatively weaker in resisting the torsional forces associated with subduing large, struggling terrestrial prey. Rayfield and colleagues' 2007 analysis found that the Baryonyx rostrum was biomechanically similar to that of the gharial and the African slender-snouted crocodile (Mecistops cataphractus), while differing substantially from the broader, more torsion-resistant skulls of generalised predatory theropods.12 Cuff and Rayfield's 2013 expanded analysis, which compared Baryonyx and Spinosaurus rostra with three extant crocodilian species, confirmed that spinosaurid snout mechanics were most consistent with piscivory, though the authors noted that Baryonyx, with its somewhat more robust rostrum, may have had a broader dietary range than the more gracile-snouted Spinosaurus.11
The forelimbs of spinosaurids are proportionally larger and more robust than those of most other large theropods, and the first digit (thumb) bears a massively enlarged, laterally compressed ungual claw. In Baryonyx, this claw measured approximately 31 centimetres along its outer curve, making it one of the largest manual claws known from any theropod.3 The functional significance of this claw has been interpreted as an adaptation for hooking and securing large fish, analogous to the fishing behaviour of modern bears, though it may also have served in defence or intraspecific combat.
The most visually striking feature of spinosaurids is the dorsal sail formed by greatly elongated neural spines on the trunk and sacral vertebrae. In Spinosaurus aegyptiacus, the longest neural spines reached heights of approximately 1.65 metres above the centra, creating a structure that would have been prominently visible in life.1, 7 The function of this sail has been debated since Stromer's original description. Proposed functions include thermoregulation (the large surface area could facilitate heat exchange with the environment), sexual display or species recognition, fat storage (analogous to camel humps, though the neural spine morphology is more consistent with a sail than a hump), and hydrodynamic stabilisation during swimming.7, 16 The neural spines vary in height across spinosaurid taxa: they are tallest in Spinosaurus, moderately elevated in Suchomimus, and relatively low in Baryonyx, suggesting that the sail became progressively more exaggerated in certain lineages over evolutionary time.
Evidence for semi-aquatic habits
The question of whether spinosaurids — and Spinosaurus in particular — were semi-aquatic animals has generated one of the most active debates in modern dinosaur palaeontology. The evidence bearing on this question comes from multiple independent sources: oxygen isotope geochemistry, bone microstructure, functional morphology, and taphonomic context.
The first quantitative evidence for semi-aquatic habits came from Amiot and colleagues' 2010 analysis of oxygen isotope ratios in spinosaurid tooth enamel. By comparing the isotopic composition of phosphate in spinosaurid teeth with that of coexisting terrestrial theropods, crocodilians, and turtles from the same localities in Africa, Europe, South America, and Asia, the authors found that spinosaurid oxygen isotope values consistently fell between those of fully terrestrial dinosaurs and those of semi-aquatic crocodilians, a pattern consistent with animals that spent a substantial fraction of their time in or near water, analogous to modern hippopotamuses or crocodilians.8
In 2014, Ibrahim and colleagues published a landmark description of a partial Spinosaurus skeleton from the Cenomanian Kem Kem beds of Morocco (designated the neotype), which revealed a suite of anatomical features interpreted as aquatic adaptations: a reduced pelvic girdle and short hindlimbs relative to body size, retracted fleshy nostrils positioned near the mid-region of the skull, and — most strikingly — limb bones that were dense and solid, lacking the open medullary cavity found in virtually all other theropod dinosaurs. This extreme bone compactness, a condition known as osteosclerosis, is a well-established adaptation for buoyancy control in secondarily aquatic tetrapods, observed in modern sirenians (manatees and dugongs), hippopotamuses, and certain aquatic birds such as penguins.7
The aquatic interpretation was dramatically reinforced in 2020 when Ibrahim and colleagues described a newly excavated tail from the same Moroccan site. The tail bore tall, elongate neural spines and broadly expanded chevrons that together created a laterally flattened, paddle-like profile unlike that of any other known theropod. Hydrodynamic modelling demonstrated that this tail could generate substantially greater thrust in water than the tails of other theropods and even some aquatic animals, suggesting that Spinosaurus was capable of tail-propelled swimming.9
In 2022, Fabbri and colleagues provided the broadest comparative test of the aquatic hypothesis by analysing bone compactness in 380 bones from a wide range of extant and extinct amniotes. Using phylogenetic flexible discriminant analysis, they found that the bone density values of Spinosaurus and Baryonyx fell squarely within the range of subaqueous foragers — animals that actively pursue prey underwater — while Suchomimus, despite being a close relative, had bone density values more consistent with non-diving wading animals.10 This result suggested ecological disparity within Spinosauridae, with some taxa (especially Spinosaurus and Baryonyx) being more aquatically adapted than others.
Bone compactness values across spinosaurids and comparative taxa10
The debate, however, is far from settled. Hone and Holtz's 2021 review cautioned that many of the features cited as aquatic adaptations in Spinosaurus are also consistent with a shoreline-wading ecology analogous to that of modern herons or storks, rather than active underwater pursuit predation. They argued that the retracted nostrils, dense bones, and reduced hindlimbs could reflect an animal that fished from shorelines and shallow water rather than one that routinely submerged and swam after prey.16 The distinction between a "wading heron" model and an "aquatic pursuit predator" model for Spinosaurus remains one of the central open questions in spinosaurid research.
Piscivorous adaptations and feeding ecology
The evidence for fish-eating in spinosaurids is among the strongest dietary evidence available for any non-avian dinosaur, resting on direct gut-content data, functional morphology, isotope geochemistry, and ecological analogy. The holotype of Baryonyx walkeri preserves acid-etched scales of the large actinopterygian fish Scheenstia mantelli and partially digested bones of a juvenile iguanodontid within the abdominal region, demonstrating that Baryonyx consumed both fish and terrestrial prey.3 This mixed dietary signal indicates that at least some spinosaurids were not obligate piscivores but rather generalist predators with a strong preference for aquatic prey.
The functional morphology of the spinosaurid skull reinforces the piscivorous interpretation. The elongated snout, terminal rosette, and conical teeth are strikingly convergent with the cranial morphology of modern longirostrine crocodilians, particularly the gharial, which feeds almost exclusively on fish. The biomechanical analyses by Rayfield and colleagues and by Cuff and Rayfield demonstrated that spinosaurid rostra were adapted for rapid lateral strikes in water, generating strong dorsoventral bending forces while minimising the torsional loading that would be imposed by large struggling prey.11, 12
An additional line of evidence for aquatic prey detection was identified by Vullo and colleagues, who in 2016 documented a striking convergence between the jaw morphology of spinosaurids and that of pike conger eels. Both groups possess elongated jaws with numerous small, conical teeth and — in the case of spinosaurids — small pits on the snout surface that have been interpreted as housing integumentary pressure receptors analogous to those in crocodilians, which detect pressure waves generated by moving prey in water.17 If confirmed, the presence of such mechanoreceptors would represent a powerful adaptation for detecting and capturing fish in murky or turbid aquatic environments, reinforcing the semi-aquatic interpretation of spinosaurid ecology.
The ecological environments in which spinosaurids are found further support their association with aquatic habitats. Spinosaurid fossils are overwhelmingly recovered from fluvial and deltaic depositional environments — river systems, floodplains, and coastal lagoons — rather than from the arid, inland environments that yield many other large theropods. The Kem Kem beds of Morocco, which have produced Spinosaurus and abundant spinosaurid teeth, represent a vast Cenomanian river system teeming with large fish, including coelacanths, lungfish, and the giant sawfish Onchopristis, whose serrated rostral denticles are frequently found alongside spinosaurid material.7, 16 The Wealden Group deposits that yielded Baryonyx similarly represent freshwater fluvial environments, consistent with a habitat association between spinosaurids and well-watered lowland ecosystems.
The rediscovery of Spinosaurus
The destruction of Stromer's holotype in 1944 left Spinosaurus aegyptiacus as one of the most poorly known of all large theropods for the remainder of the twentieth century. Although fragmentary spinosaurid teeth and bones continued to be reported from the Cretaceous of North Africa, no substantial new material clearly attributable to Spinosaurus was described until the twenty-first century, and the animal's size, proportions, and ecology remained largely conjectural.
This changed dramatically in 2014 with the publication of a partial skeleton from the Cenomanian Kem Kem beds of southeastern Morocco by Nizar Ibrahim and colleagues. The specimen, designated the neotype of Spinosaurus aegyptiacus, included parts of the skull, vertebral column, pelvis, and hindlimb, and its analysis revealed an animal fundamentally unlike the reconstruction that had been assumed for nearly a century. Rather than a conventional large theropod with an unusually tall sail, Spinosaurus emerged as a bizarre, specialised predator with proportionally tiny hindlimbs, a barrel-shaped torso, dense solid bones, and a centre of mass positioned far forward of the hip joint — a body plan that Ibrahim and colleagues interpreted as incompatible with sustained bipedal terrestrial locomotion and instead adapted for a semi-aquatic, quadrupedally-assisted mode of life.7
The 2020 description of the paddle-shaped tail from the same Moroccan locality added another extraordinary dimension to the Spinosaurus reconstruction. The preserved caudal vertebrae bore neural spines and chevrons that were elongated and unexpanded, creating a fin-like tail profile unmatched by any other known theropod. Experimental modelling using a flexible robotic tail demonstrated that this morphology generated eight times more thrust in water than the tails of other theropods such as Coelophysis and Allosaurus.9
Global distribution
Spinosaurids achieved a remarkably wide geographic distribution across both Gondwanan and Laurasian landmasses during the Cretaceous. The group's fossil record spans every major continental region except Antarctica and Australia, though sampling biases in those areas make their true absence uncertain. Understanding spinosaurid biogeography requires consideration of the palaeogeographic configuration of the Cretaceous world, in which Gondwana was fragmenting and intermittent land bridges or shallow seaways permitted periodic faunal interchange between continental blocks.
Europe has yielded some of the earliest and most diverse spinosaurid material. Baryonyx walkeri from the Barremian of England (approximately 130–125 million years ago) is among the oldest well-known spinosaurids. The Isle of Wight has produced the recently described Ceratosuchops inferodios and Riparovenator milnerae, both from the Wessex Formation (Barremian), demonstrating that multiple spinosaurid species coexisted in the same region during the Early Cretaceous.13 Additional spinosaurid material is known from the Early Cretaceous of Spain and Portugal, confirming a broad European distribution during the Barremian and Aptian stages.
Africa has produced the most spectacular spinosaurid material, including both Suchomimus tenerensis from the Aptian of Niger and Spinosaurus aegyptiacus from the Cenomanian of Egypt and Morocco. The Kem Kem beds of Morocco are particularly rich, having yielded abundant isolated spinosaurid teeth alongside the neotype skeleton described in 2014.4, 7 South America preserves several spinosaurine taxa from the Early Cretaceous of Brazil, most notably Irritator challengeri from the Albian Santana Formation, known from the most complete spinosaurid skull discovered to date, and Oxalaia quilombensis from the Cenomanian Alcântara Formation.15
Asia was long considered devoid of definitive spinosaurid fossils until Allain and colleagues described Ichthyovenator laosensis from the Aptian Grès supérieurs Formation of Laos in 2012 — the first unambiguous spinosaurid from the Asian continent. Ichthyovenator is notable for its distinctive neural spine morphology, in which the sail appears to have had a sinusoidal profile with a gap or notch between the dorsal and sacral sail regions, unlike the continuous sail of Spinosaurus.14 Isolated spinosaurid teeth have also been reported from the Early Cretaceous of Thailand, Japan, and China, suggesting that the family may have been more widespread across Asia than the skeletal record alone indicates.
Major spinosaurid taxa and their geographic and temporal distribution3, 4, 7, 13, 14, 15
| Taxon | Subfamily | Region | Age (stage) | Estimated length |
|---|---|---|---|---|
| Baryonyx walkeri | Baryonychinae | England | Barremian | ~7.5–10 m |
| Ceratosuchops inferodios | Baryonychinae | England | Barremian | ~8 m |
| Riparovenator milnerae | Baryonychinae | England | Barremian | ~8 m |
| Suchomimus tenerensis | Baryonychinae | Niger | Aptian | ~11 m |
| Ichthyovenator laosensis | Spinosaurinae | Laos | Aptian | ~8.5 m |
| Irritator challengeri | Spinosaurinae | Brazil | Albian | ~6–8 m |
| Spinosaurus aegyptiacus | Spinosaurinae | Egypt, Morocco | Cenomanian | ~14–15 m |
Ecology and habitat associations
The depositional environments from which spinosaurid fossils are recovered paint a consistent picture of animals closely associated with freshwater and marginal-marine ecosystems. Spinosaurids are found overwhelmingly in fluvial sediments — the deposits of ancient river channels, floodplains, and deltaic settings — rather than in the aeolian (wind-deposited) or lacustrine sediments that produce many other large theropods. This association is not an artefact of preservation bias; other theropod taxa from the same formations, such as the carcharodontosaurids and abelisaurids that coexisted with Spinosaurus in the Kem Kem beds, do not show the same preferential association with fluvial facies.7, 16
The Kem Kem ecosystem of mid-Cretaceous North Africa provides the richest ecological context for understanding spinosaurid palaeoecology. During the Cenomanian (approximately 100–94 million years ago), the region that is now southeastern Morocco was a vast, low-lying river system bordered by extensive wetlands, with a warm, humid climate supporting lush vegetation and an extraordinarily diverse aquatic fauna. The fish assemblage included coelacanths (Mawsonia), lungfish (ceratodus-like forms), polypterids, large teleosts, and the giant sawfish Onchopristis numidus, some of which reached body lengths exceeding three metres. This abundance of large aquatic prey would have provided a rich food base for a large piscivorous predator, and the co-occurrence of multiple large theropod taxa in the same ecosystem — including Spinosaurus, Carcharodontosaurus, and the abelisaurid Deltadromeus — has been interpreted as evidence of niche partitioning, with Spinosaurus exploiting aquatic resources that the more conventionally terrestrial theropods could not efficiently access.7, 16
Whether spinosaurids interacted with marine environments remains less clear. The majority of spinosaurid localities are interpreted as freshwater systems. However, some Kem Kem deposits preserve evidence of tidal influence and occasional marine incursions, and spinosaurid teeth have occasionally been found in near-coastal or shallow-marine sediments, suggesting that some spinosaurids may have exploited estuarine or lagoonal environments, much as modern saltwater crocodiles utilise both freshwater and marine habitats.8
Ongoing debates and recent discoveries
Spinosauridae is one of the most actively debated dinosaur groups in contemporary palaeontology, with several major questions remaining unresolved despite rapid progress in recent years. The most prominent debate concerns the degree of aquatic adaptation in Spinosaurus. The Ibrahim-led research programme has proposed a model of Spinosaurus as an active, tail-propelled pursuit predator that routinely dived and swam after fish underwater, drawing on the evidence of the paddle tail, osteosclerotic bone, and reduced hindlimbs.7, 9 Against this, Hone and Holtz have argued for a more conservative "wading" or "shoreline generalist" model, in which Spinosaurus fished primarily from riverbanks and shallow water, wading rather than swimming, in a manner analogous to large herons or storks. They note that no known Spinosaurus specimen preserves webbed feet or flippers, that the forelimbs retained functional grasping claws unsuited to aquatic propulsion, and that the sail would have imposed significant hydrodynamic drag during submerged swimming.16
The Fabbri et al. 2022 bone density study partially resolved this debate by demonstrating that different spinosaurid species adopted different degrees of aquatic commitment, with Spinosaurus and Baryonyx showing evidence of subaqueous foraging while Suchomimus appears to have been a non-diving wader.10 This result is consistent with the emerging picture of Spinosauridae as a family that explored a spectrum of semi-aquatic ecologies rather than a single stereotyped lifestyle.
The taxonomy of Spinosaurus itself continues to evolve. The relationship between Spinosaurus aegyptiacus from Egypt, the Moroccan Kem Kem material described by Ibrahim and colleagues, and the putative separate genus Sigilmassasaurus (erected on the basis of isolated vertebrae from Morocco) has been debated, with some authors treating all North African spinosaurine material as a single species and others arguing for the recognition of multiple distinct taxa.15 The question of how many Spinosaurus species existed across the vast river systems of mid-Cretaceous North Africa remains unresolved.
The discovery of new spinosaurid taxa continues at a remarkable pace. In 2021 alone, two new genera were described — Ceratosuchops and Riparovenator — from the Isle of Wight.13 Each new discovery refines the picture of spinosaurid anatomy, phylogeny, and ecology, while simultaneously opening new questions about the diversity and evolutionary success of these uniquely adapted theropod dinosaurs. Spinosauridae, once regarded as an evolutionary footnote, is now recognised as a major radiation of large predatory dinosaurs that evolved one of the most distinctive ecological strategies in the 165-million-year history of the Mesozoic dinosaur dynasty.
References
Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharîje-Stufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen., nov. spec.
New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae
The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the Early Cretaceous of Laos
Evaluating the ecology of Spinosaurus: shoreline generalist or aquatic pursuit specialist?
Convergence between the jaws of pike conger eels and spinosaurid dinosaurs reveals a trophic specialisation in early spinosaurids