Overview
- The founder effect occurs when a small number of individuals from a larger population establish a new colony, carrying only a fraction of the original population's genetic diversity and producing allele frequencies that may differ dramatically from the source group.
- Classic examples include the high prevalence of Ellis-van Creveld syndrome among the Old Order Amish, variegate porphyria among Afrikaners, achromatopsia on Pingelap atoll, and multiple recessive disease alleles in Quebec founder populations — all traceable to chance sampling in small founding groups.
- Ernst Mayr proposed that founder events could trigger rapid speciation by reorganizing epistatic gene complexes in small peripheral isolates, a model that remains influential in island biogeography and invasion biology despite ongoing theoretical debate.
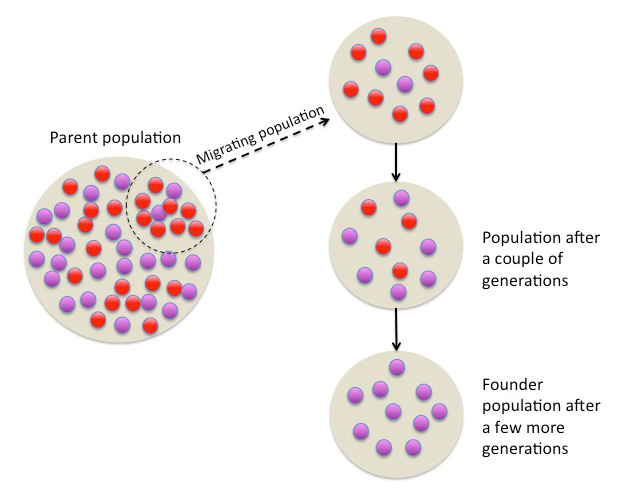
The founder effect is a special case of genetic drift in which a new population is established by a small number of individuals drawn from a larger source population. Because the founders represent only a limited sample of the parent population's gene pool, the new colony typically begins with allele frequencies that differ from those of the source, and it may lack rare alleles entirely or carry otherwise uncommon variants at elevated frequency. The magnitude of genetic change depends on the number of founders, the degree to which their genotypes are representative of the source population, and whether the colony remains genetically isolated during its early growth. First described conceptually by Ernst Mayr in 1954, the founder effect has been invoked to explain patterns of genetic disease in human isolates, the genetics of island colonisation, and the role of small populations in triggering speciation.1, 8
{kind=link}
Definition and mechanism
The founder effect arises from the same stochastic sampling process that underlies all genetic drift: in a finite population, the alleles transmitted to the next generation are a random sample of the parental alleles, and smaller samples deviate more from the population mean than larger ones. What distinguishes the founder effect from ordinary drift operating within a continuously existing population is the abruptness and severity of the sampling event. When a handful of colonists establish a new population, the effective population size at the founding moment may be as small as a single mating pair, and the allele frequencies in the nascent colony are determined by whatever genetic variation those few individuals happen to carry.8, 9
The mathematical framework for understanding the founder effect builds directly on Sewall Wright's theory of genetic drift in finite populations. Wright showed in 1931 that the variance in allele frequency change per generation for a neutral allele at frequency p is approximately p(1 − p)/(2Ne), where Ne is the effective population size.18 During a founding event, Ne is extremely small, and the sampling variance is correspondingly enormous. Nei, Maruyama, and Chakraborty formalised the genetic consequences in 1975, demonstrating that a severe reduction in population size rapidly eliminates rare alleles and reduces heterozygosity, and that while heterozygosity recovers relatively quickly once the population expands, the full spectrum of allelic diversity takes far longer to be restored through new mutation.3
The expected reduction in heterozygosity following a bottleneck of Ne founders lasting one generation is approximately 1/(2Ne). For a founding group of 10 individuals, this represents a 5 percent loss of heterozygosity in a single generation, but the loss of rare alleles is far more severe because low-frequency variants are the most likely to be absent from a small sample.3, 12 If the population remains small for multiple generations before expanding, the cumulative effect is amplified considerably, because the long-term effective population size is determined by the harmonic mean of population sizes across generations, which is dominated by the smallest values.12
Distinction from the bottleneck effect
The founder effect and the population bottleneck are closely related phenomena that both involve drastic reductions in population size, but they differ in their ecological context. A bottleneck occurs when an existing population is reduced to a small fraction of its former size by some catastrophic event, such as a disease epidemic, environmental disaster, or overhunting, after which the surviving remnant continues to occupy the same geographic range. The founder effect, by contrast, involves the geographic separation of a small group from the parent population to establish a new colony in a previously unoccupied area.8
The genetic consequences of the two processes are qualitatively similar: both result in the loss of rare alleles, reduced heterozygosity, and the potential elevation of otherwise uncommon variants to high frequency through chance. The key distinction is that a bottleneck temporarily reduces the effective population size of a population that subsequently recovers in the same location, whereas a founder event creates a new, geographically isolated population whose subsequent evolutionary trajectory may diverge from that of the source. This geographic isolation is critical because it prevents gene flow from the source population, which would otherwise tend to restore the original allele frequencies. The combination of small founding size and reproductive isolation makes the founder effect a potent mechanism for generating genetic divergence between populations.3, 8, 15
Classic examples in human populations
Some of the most thoroughly documented examples of the founder effect involve human populations that trace their ancestry to small, well-characterised founding groups. The Old Order Amish of Lancaster County, Pennsylvania, descend from approximately 200 Swiss-German immigrants who arrived in the early eighteenth century. Victor McKusick's pioneering medical genetics studies of the Amish revealed that Ellis-van Creveld syndrome, a recessive condition characterised by short-limbed dwarfism, polydactyly, and congenital heart defects, occurs at a frequency roughly a hundredfold higher among the Lancaster County Amish than in the general population. The elevated prevalence is traceable to a single carrier couple among the original founders, and the community's small size, endogamy, and rapid growth through high birth rates rather than immigration have maintained the allele at high frequency ever since.4
The Afrikaner population of South Africa provides an equally striking illustration. Much of the Afrikaner gene pool derives from a small number of Dutch, German, and French Huguenot settlers who arrived at the Cape of Good Hope in the seventeenth century. Among these founders, at least one individual carried the R59W mutation in the protoporphyrinogen oxidase gene, which causes variegate porphyria, a disorder of haem biosynthesis that produces photosensitive skin lesions and episodes of acute abdominal pain. Meissner and colleagues demonstrated in 1996 that approximately 95 percent of South African variegate porphyria cases carry this single founder mutation, which has reached a carrier frequency of roughly 1 in 300 among Afrikaners, orders of magnitude higher than in any other population.5
On the remote Pacific atoll of Pingelap in Micronesia, a devastating typhoon around 1775 reduced the population to approximately 20 survivors. Among these survivors, at least one individual carried a recessive mutation in the CNGB3 gene that causes achromatopsia, or complete colour blindness. Because the post-typhoon population was extremely small and geographically isolated, the allele drifted to high frequency during the subsequent demographic recovery. Sundin and colleagues reported in 2000 that achromatopsia affects between 4 and 10 percent of the Pingelapese population, roughly a thousandfold higher than the global prevalence, making it one of the most dramatic examples of a single founder event shaping the disease burden of an entire community for centuries.6
The Ashkenazi Jewish population experienced a well-documented contraction during the medieval period, when the effective founding population has been estimated at roughly 350 to 500 individuals. Risch and colleagues analysed the geographic distribution of multiple recessive disease alleles among Ashkenazi Jews, including those responsible for Tay-Sachs disease, Gaucher disease, and familial dysautonomia, and concluded that the pattern of elevated carrier frequencies is more consistent with genetic drift during founder events than with heterozygote advantage driven by natural selection.7 Ostrer confirmed that many of the disease-associated mutations found at elevated frequency in Ashkenazi populations can be traced to a small number of ancestral haplotypes, consistent with the expansion of founder alleles rather than recurrent mutation or balancing selection.13
The Quebec founder population
The French-Canadian population of Quebec represents one of the best-characterised founder populations in the world, owing to the availability of detailed parish records extending back to the seventeenth century. The colony was established by approximately 8,500 French immigrants who arrived between 1608 and 1759, and subsequent growth was driven almost entirely by natural increase rather than continued immigration. Laberge and colleagues demonstrated that the genetic structure of modern French Canadians reflects this founding history, with reduced genetic diversity relative to the French source population and elevated frequencies of several recessive disease alleles that are rare or absent in France.14
Several genetic diseases occur at unusually high frequency in specific regions of Quebec as a direct consequence of local founder effects within the broader French-Canadian population. Hereditary tyrosinemia type I, which causes severe liver disease, reaches a carrier frequency of approximately 1 in 20 in the Saguenay-Lac-Saint-Jean region, compared with roughly 1 in 100,000 in most other populations. This extreme enrichment is attributable to the region's settlement by a small number of founding families from the broader Quebec population, creating a founder effect within a founder effect. The Quebec example demonstrates that founder effects can operate at multiple nested scales, with successive colonisation events progressively narrowing the genetic base of each daughter population.14
Serial founder effects and human migration
The concept of serial founder effects has been invoked to explain one of the most striking patterns in human population genetics: the systematic decline in genetic diversity with increasing geographic distance from sub-Saharan Africa. Ramachandran and colleagues demonstrated in 2005 that a strong linear relationship exists between a population's distance from East Africa, measured along plausible overland migration routes, and its level of microsatellite heterozygosity. Populations in sub-Saharan Africa harbour the greatest genetic diversity of any human group, while populations at the geographic extremes of the human range, such as indigenous peoples of South America and Oceania, exhibit the lowest.10
Prugnolle, Manica, and Balloux independently confirmed this gradient, showing that geographic distance from Africa is the single best predictor of neutral genetic diversity across worldwide human populations.11 The serial founder effect model proposes that as modern humans expanded out of Africa beginning roughly 50,000 to 70,000 years ago, each successive colonisation of new territory was accomplished by a small group splitting from the wavefront population. Each split constituted a founder event that randomly eliminated a portion of the genetic variation carried forward, producing a cumulative ratchet-like decline in diversity with distance from the ancestral homeland.10, 20
Excoffier, Foll, and Petit explored the broader consequences of serial founder effects during range expansions, demonstrating through simulation that the successive bottlenecks at the expansion front can produce not only reduced neutral diversity but also the random fixation of deleterious alleles through a process they termed "allele surfing." In this phenomenon, alleles present at the expansion wavefront, regardless of their fitness effects, can ride the wave of population growth to reach high frequency or fixation in newly colonised territory simply because they were present in the right place at the right time.21
Role in speciation
Ernst Mayr proposed in 1954 that founder events could play a central role in the formation of new species through a process he termed the "genetic revolution." Mayr argued that when a small founder population becomes geographically isolated from the parent species, the drastic change in allele frequencies caused by the sampling event disrupts the co-adapted gene complexes that had been maintained by stabilising selection in the large ancestral population. This disruption shifts the population to a new "adaptive peak" in Wright's fitness landscape, potentially producing rapid morphological and behavioural divergence that leads to reproductive isolation.1, 2
Mayr developed this idea more fully in his 1963 monograph Animal Species and Evolution, where he termed the process peripatric speciation, referring to speciation in small, geographically peripheral populations. He argued that the combination of strong drift in the founding population, altered selection pressures in the new environment, and the absence of gene flow from the parent population creates conditions uniquely favourable for rapid evolutionary divergence. The Hawaiian archipelago, with its spectacular radiations of honeycreepers, Drosophila, and silversword plants, was frequently cited by Mayr as exemplifying the role of founder events in generating species diversity on oceanic islands.2, 15
The theoretical basis for Mayr's genetic revolution has been debated extensively. Coyne and Orr reviewed the evidence and theoretical models in their 2004 synthesis and concluded that while founder events can certainly alter allele frequencies, the claim that such changes routinely trigger the reorganisation of co-adapted gene complexes and produce reproductive isolation remains theoretically contentious. Population genetic models suggest that founder events alone, without subsequent divergent natural selection, are unlikely to produce the kind of epistatic changes Mayr envisioned, because random drift in a small population is as likely to break down incipient reproductive barriers as to build them up.15 Nevertheless, the observation that many island radiations and cases of rapid speciation are associated with geographic colonisation by small founding groups ensures that the founder effect remains a central concept in discussions of speciation mechanisms.2, 15
Founder effects in invasive species
The introduction of non-native species to new environments frequently involves founder events, because colonising populations are typically established by a small number of individuals transported, deliberately or accidentally, from the species' native range. This creates a paradox: invasive species are often spectacularly successful in their new environments despite having passed through a genetic bottleneck that should, in theory, reduce their adaptive potential. Lee reviewed the evolutionary genetics of invasive species and noted that many successful invaders show significantly reduced genetic diversity relative to their source populations, consistent with strong founder effects during introduction.16
Several mechanisms have been proposed to resolve this "genetic paradox of invasions." Multiple independent introductions from different source populations can increase genetic diversity through admixture, sometimes generating novel genotypic combinations not found in any single source population. Rapid adaptive evolution from standing genetic variation, even if reduced, can allow populations to respond to new selective pressures. Facon and colleagues argued that inbreeding-by-environment interactions and the purging of deleterious recessive alleles during the bottleneck phase may actually facilitate adaptation in some cases, as the founder event exposes recessive alleles to selection more efficiently in homozygous individuals.17
The zebra mussel (Dreissena polymorpha), the European starling (Sturnus vulgaris), and the cane toad (Rhinella marina) all exhibit reduced genetic diversity in their introduced ranges relative to their native ranges, consistent with founder effects during colonisation. Yet in each case, the introduced populations have expanded explosively, demonstrating that the loss of genetic diversity associated with a founder event does not necessarily prevent ecological success in the short term, even if it may constrain long-term adaptive potential.16, 17
Island colonisation and genetic divergence
Oceanic islands have long served as natural laboratories for studying the founder effect because their biota must, by definition, have arrived via long-distance dispersal by a small number of colonists. The genetic diversity of island populations is almost invariably lower than that of their mainland relatives, and the magnitude of the reduction correlates with island remoteness, island area, and estimated time since colonisation. Darwin's finches of the Galápagos, Hawaiian Drosophila, and Canary Island lacertid lizards all show patterns of reduced diversity and elevated genetic differentiation among island populations that are consistent with founder effects followed by drift and local adaptation in isolation.2, 15
The founder effect on islands extends beyond simple allele frequency changes. When a founder population is small enough, the effective recombination landscape can change: linkage disequilibrium between loci is elevated because the few founder chromosomes establish a limited number of haplotype combinations, and this disequilibrium decays only slowly if the population remains small. The result is that natural selection acting on any one locus can drag linked variants along, amplifying the hitchhiking effect and further distorting allele frequencies from neutral expectations. This interaction between founder effects, linkage, and selection in small island populations may accelerate phenotypic divergence and contribute to the rapid morphological evolution often observed in island radiations.12, 15
Significance
The founder effect demonstrates that chance plays an irreducible role in evolution, particularly at the moments when new populations are established. The allele frequencies of a founding group are determined not by the adaptive value of different genotypes but by the accident of which individuals happen to colonise a new territory. This stochastic element means that populations derived from the same source can diverge genetically and phenotypically even in the absence of different selective pressures, simply because different founders carried different subsets of the ancestral gene pool.8, 9
In human genetics, the founder effect has profound practical implications. The elevated frequency of specific disease alleles in founder populations enables targeted genetic screening programmes, such as those developed for Tay-Sachs disease in Ashkenazi Jewish communities, which have dramatically reduced the incidence of affected births.7, 13 In conservation biology, understanding founder effects informs strategies for captive breeding and reintroduction programmes, where the genetic diversity of the founding stock determines the long-term viability of the new population.3 And in evolutionary biology more broadly, the founder effect remains central to debates about the relative importance of drift and selection in speciation, the genetics of island radiations, and the evolutionary dynamics of biological invasions.1, 2, 15
References
A R59W mutation in human protoporphyrinogen oxidase results in decreased enzyme activity and is prevalent in South Africans with variegate porphyria
Geographic distribution of disease mutations in the Ashkenazi Jewish population supports genetic drift over selection
Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa
The genetic structure of the French-Canadian population: evidence from autosomal, Y-chromosomal, and mitochondrial DNA
The genetic paradox of invasions revisited: the potential role of inbreeding x environment interactions in invasion success
Genetic heritage of the Balto-Slavic speaking populations: a synthesis of autosomal, mitochondrial and Y-chromosomal data