Overview
- A ring species is a chain of interbreeding populations that encircles a geographic barrier, where the terminal populations at the ends of the chain overlap but cannot interbreed, providing a spatial snapshot of speciation as a gradual, continuous process.
- The Ensatina salamanders of California's Central Valley and the greenish warbler complex around the Himalayas are the most thoroughly studied examples, each demonstrating how cumulative genetic and phenotypic divergence along a geographic arc can produce reproductive isolation without any sharp boundary between species.
- Ring species are exceptionally rare in nature and several classic examples, including the herring gull complex, have been reinterpreted by molecular phylogenetics, yet the concept remains a powerful illustration of how geographic distance, reduced gene flow, and gradual divergence drive the origin of new species.
A ring species is a connected series of neighbouring populations that encircles a geographic barrier, where each adjacent population can interbreed with its neighbours, yet the terminal populations at the two ends of the chain overlap geographically and are reproductively isolated from one another. The concept, first articulated by Ernst Mayr in the mid-twentieth century, represents one of the most striking illustrations of how speciation can occur as a gradual, continuous process rather than through an abrupt split.14, 15 In a ring species, one can trace the transition from a single interbreeding population to two distinct species along a spatial continuum, with no clear point at which one species ends and the next begins. This spatial gradient of divergence effectively lays out in geography what ordinarily unfolds invisibly in time, offering a rare living snapshot of speciation in progress.2, 3
The concept and its history
The idea of a ring species emerged from the broader study of geographic variation in the early twentieth century. Ernst Mayr, in his foundational work on geographic speciation, described the phenomenon as a Rassenkreis (German for "circle of races") in which a chain of subspecies forms a ring around a barrier, with the terminal forms behaving as distinct species where they meet.14, 15 Mayr considered ring species to be the most compelling demonstration of his theory that geographic isolation and gradual divergence, rather than sudden genetic revolutions, are the primary drivers of speciation. The concept directly supports the model of speciation by distance: as populations spread around a barrier, gene flow between distant populations diminishes, allowing independent genetic and phenotypic divergence to accumulate along each arm of the expanding range.2, 16
{kind=link}
Sewall Wright's mathematical framework of isolation by distance provided the theoretical foundation for understanding how gene flow declines with geographic separation even in continuously distributed populations.16 In a ring species, this decline is made visible: adjacent populations exchange genes freely, but the cumulative reduction in gene flow around the entire circuit is sufficient to produce reproductive isolation between the terminal populations. The ring species concept thus bridges microevolution (the small changes between neighbouring populations that any biologist can observe) and macroevolution (the origin of new species), demonstrating that speciation can be the endpoint of a continuous process of gradual divergence.2, 3
Ensatina salamanders of California
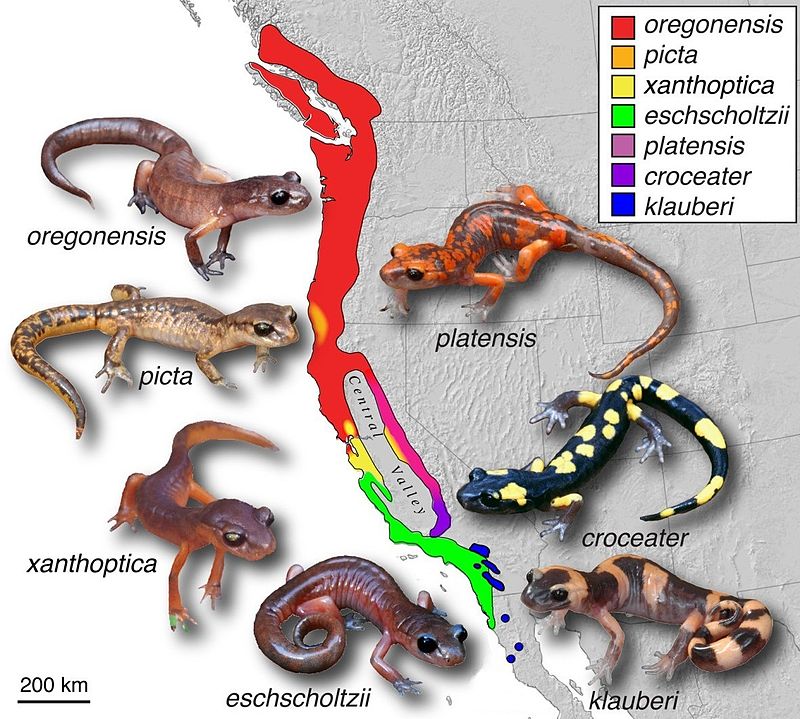
The most thoroughly documented example of a ring species is the Ensatina eschscholtzii complex, a group of plethodontid (lungless) salamanders distributed around the Central Valley of California. The complex was first proposed as a ring species by Robert Stebbins in the 1940s and has been the subject of intensive morphological, ecological, and molecular investigation for more than half a century, particularly through the work of David Wake and his collaborators at the University of California, Berkeley.4, 5

{kind=link}
The geographic pattern is elegant. Ensatina populations form a roughly horseshoe-shaped distribution along the mountain ranges flanking California's Central Valley, which is itself too hot and dry to support the salamanders. From a presumed ancestral population in the mountains of northern California and southern Oregon, two arms of populations extend southward: one along the Coast Ranges on the west side of the valley and another along the Sierra Nevada on the east side. Along each arm, adjacent populations intergrade freely, showing clinal variation in colour pattern, body size, and allozyme frequencies.4, 5 However, at the southern end of the valley in the mountains of southern California, the two arms come into contact, and here the coastal form (E. e. eschscholtzii, a relatively plain brownish-orange salamander) and the inland form (E. e. klauberi, a boldly blotched black-and-yellow mimic of the toxic newt Taricha) coexist with little or no hybridisation, behaving as reproductively isolated species.4, 6
Molecular phylogeographic studies using mitochondrial DNA have confirmed the overall pattern of divergence predicted by the ring species model, showing a progressive increase in genetic distance with geographic distance around the ring.6, 20 However, these same studies have revealed complexities that complicate the simplest version of the ring species narrative. In particular, the phylogenetic relationships among the subspecies do not form a simple bifurcating tree consistent with a single expansion around the valley; instead, some lineages appear to have experienced periods of isolation and secondary contact, suggesting that the history of Ensatina involves both continuous gene flow along the ring and episodes of fragmentation caused by Pleistocene glacial cycles and geological changes.7, 20
The question of hybridisation at contact zones within the ring has received particular attention. While the terminal overlap in southern California shows strong reproductive isolation, several intermediate contact zones along the ring display varying degrees of hybridisation and intergradation. Some contacts between subspecies show narrow but stable hybrid zones, while others show broad intergradation. This mosaic of outcomes along a single geographic circuit demonstrates that reproductive isolation does not accumulate uniformly but depends on the specific traits diverging in each lineage and the ecological context of each contact zone.4, 21 Wake has argued that this complexity, far from undermining the ring species concept, actually makes Ensatina a richer model for understanding speciation as a multidimensional process involving the independent divergence of multiple traits in different parts of the range.4
The greenish warbler complex
The greenish warbler (Phylloscopus trochiloides) complex, which encircles the Tibetan Plateau and the western Himalayas, is the best-supported avian ring species and the example for which quantitative data on gradual divergence is most complete.

_-_48728300083.jpg){kind=link}
The complex was identified as a ring species through a combination of morphometric analysis, song recordings, playback experiments, and molecular phylogenetics, primarily through the work of Darren Irwin and Trevor Price.1, 8, 9
Populations of greenish warblers form a ring of breeding ranges around the high, barren Tibetan Plateau, which serves as the uninhabitable central barrier. From an inferred ancestral range in the southern Himalayas, two chains of populations extend northward: a western chain through Afghanistan, Tajikistan, and Kazakhstan, and an eastern chain through China and Mongolia. Along each arm, neighbouring populations interbreed and show gradual, clinal changes in plumage colour, body size, wing-bar pattern, and song characteristics.1, 9 At the northern end of the ring, in central Siberia, the western form (P. t. viridanus) and the eastern form (P. t. plumbeitarsus) come into contact in a zone of sympatry. Here, the two forms differ markedly in song, respond strongly to playback of their own song type but weakly or not at all to the other form's song, and do not hybridise to any significant extent, behaving as reproductively distinct species.8, 10
Irwin and colleagues demonstrated that the degree of genetic divergence between populations increases predictably with the geographic distance measured around the ring rather than the straight-line distance between them, confirming that the barrier (the Tibetan Plateau) has forced gene flow to follow the circuitous route around it.1 Song divergence follows a parallel gradient, with song characteristics changing gradually along each arm but differing dramatically between the two terminal forms. Because song is the primary mechanism of mate recognition in Phylloscopus warblers, the gradual divergence in song along the ring provides a direct mechanistic link between the accumulation of small differences and the emergence of reproductive isolation.8, 10
More recent genomic analyses have added nuance to the greenish warbler ring, as they have for Ensatina. Some studies suggest that gene flow has not been perfectly continuous around the ring and that certain segments may have experienced periods of reduced connectivity, particularly during glacial periods when suitable habitat contracted.18 Nevertheless, the overall pattern of gradual, distance-dependent divergence culminating in reproductive isolation at the point of terminal overlap remains robust, and the greenish warbler complex continues to be regarded as the clearest example of speciation by distance in a ring configuration.1, 2
The herring gull complex and its reinterpretation
For much of the twentieth century, the herring gull (Larus argentatus) and lesser black-backed gull (Larus fuscus) complex was presented as the textbook example of a ring species in ornithology and was featured prominently in introductory biology courses. The narrative was appealing: a chain of gull populations was said to encircle the Arctic, with each adjacent population interbreeding freely and showing gradual variation in plumage and size, until the chain completed its circuit in northwestern Europe, where herring gulls and lesser black-backed gulls breed side by side as distinct, non-interbreeding species.14, 15
Molecular phylogenetic studies beginning in the late 1990s and early 2000s challenged this interpretation. Analyses of mitochondrial DNA and microsatellite markers by Dorit Liebers, Andreas Helbig, and colleagues revealed that the evolutionary history of the Larus argentatus/L. fuscus complex is far more complicated than a simple ring of gradually diverging populations.12 Rather than a single continuous expansion around the Arctic, the molecular evidence suggests multiple independent colonisation events, periods of geographic isolation in separate glacial refugia, and episodes of secondary contact and hybridisation. The relationships among the various gull taxa around the circumpolar ring do not form the pattern of stepwise divergence predicted by the ring species model; instead, some geographically distant populations are genetically more similar to each other than to their immediate neighbours, indicating independent origins or extensive long-distance gene flow.11, 13
Crochet and colleagues independently arrived at similar conclusions, finding that the phylogeographic structure of the complex reflected a history of fragmentation and recolonisation from multiple refugia during Pleistocene glaciations rather than a single wave of expansion around the Arctic.13 The gull complex is now widely regarded as a case of repeated allopatric divergence and secondary contact rather than a true ring species, though the details of its evolutionary history remain debated. The reinterpretation of the herring gull complex illustrates both the power of molecular phylogenetics to test long-standing biogeographic hypotheses and the difficulty of distinguishing ring-species dynamics from other modes of speciation in nature.11, 12, 13
Gene flow gradients and isolation by distance
The mechanism underlying ring species is fundamentally one of declining gene flow with geographic distance. In any continuously distributed species, individuals are more likely to mate with nearby individuals than with distant ones, and the genetic consequences of this spatial structure were formalised by Sewall Wright in his isolation-by-distance model.16 Under isolation by distance, the correlation in allele frequencies between two populations decays as a function of the geographic distance between them, even in the absence of any physical barrier to dispersal. In most species, this distance-dependent differentiation produces modest clinal variation but does not lead to speciation, because gene flow across the entire range, even if indirect and mediated through a chain of intermediate populations, is sufficient to maintain cohesion.16, 19
What makes a ring species exceptional is the presence of a central barrier that prevents direct gene flow between populations on opposite sides, forcing all genetic exchange to travel the long way around the ring. This effectively doubles the geographic distance between the two arms of the population relative to what it would be without the barrier, pushing the most distant populations past the threshold at which accumulated genetic and phenotypic differences produce reproductive isolation.2, 3 The ring species concept therefore predicts that reproductive isolation should be correlated with the distance measured around the ring (the path distance) rather than the straight-line (Euclidean) distance, a prediction that has been confirmed in both the greenish warbler and Ensatina systems.1, 6
The specific traits that diverge along the gene flow gradient vary among ring species and reflect the different selective pressures and drift effects operating in each arm of the ring. In greenish warblers, song divergence appears to be the primary driver of reproductive isolation at the zone of terminal overlap, with natural selection on foraging ecology and sexual selection on song both contributing to differentiation along each arm.8, 10 In Ensatina, colour pattern divergence, possibly driven by mimicry of toxic newts in some populations, plays a central role, along with genetic incompatibilities revealed by reduced hybrid fitness in certain contact zones.4, 21 In both cases, the ring provides a natural experiment in which the same ancestral gene pool has been subdivided and subjected to different evolutionary forces along two independent pathways, with the zone of terminal overlap serving as the test of whether those forces have been sufficient to produce speciation.2
Rarity and complications
True ring species are exceedingly rare in nature. Despite the conceptual elegance of the model, very few biological systems meet all of the criteria simultaneously: continuous distribution around a barrier, gradual clinal variation along each arm, gene flow between all adjacent populations, and reproductive isolation at the point of terminal overlap. Coyne and Orr, in their comprehensive review of speciation, note that fewer than a handful of convincing cases have been documented, and even these are subject to ongoing debate as molecular data reveal historical complexities that deviate from the idealised ring model.3
Several factors make perfect ring species unlikely. The geographic and ecological conditions required — a barrier of the right size and shape, surrounded by continuously habitable terrain — are themselves uncommon. Moreover, the process of range expansion around a barrier must proceed without interruption by climatic or geological events that fragment populations and break the chain of gene flow, a condition that is rarely met over the timescales (thousands to millions of years) required for sufficient divergence to accumulate.2, 19 Pleistocene glacial cycles, in particular, have repeatedly fragmented and reconnected populations across the Northern Hemisphere, superimposing episodes of allopatric divergence on any pre-existing ring-species dynamics, as seen in both the Ensatina and gull systems.7, 11
The complications revealed by molecular studies have led some biologists to question whether any real system perfectly matches the theoretical ideal of a ring species. However, others have argued that the value of the ring species concept does not depend on the existence of perfect examples. Even imperfect rings, in which the history includes some periods of fragmentation and secondary contact, demonstrate the fundamental principle that reproductive isolation can arise through the gradual accumulation of differences along a geographic continuum. The question is not whether ring species exist in a pure form but whether the spatial pattern of divergence observed in these systems is informative about the process of speciation, and on that point the evidence from Ensatina and the greenish warbler complex is compelling.2, 4
Genetic divergence around the Ensatina ring6, 20
Significance for understanding speciation
Ring species occupy a unique place in evolutionary biology because they make the process of speciation visible in space. Speciation is ordinarily inferred from its results — the existence of distinct, reproductively isolated species — and the intermediate stages, during which populations are partially differentiated but not yet fully isolated, are difficult to observe because they are transient in time. A ring species, by contrast, preserves all of these intermediate stages simultaneously: fully compatible populations at the starting point, progressively differentiated populations along each arm, and reproductively isolated forms at the terminal overlap. Walking along the ring is, in a sense, walking through the process of speciation.2, 3
This spatial representation of a temporal process has several important implications. It demonstrates that species boundaries need not be sharp and that the transition from one species to two can be a continuous gradient rather than a discrete event. It shows that reproductive isolation can evolve as a byproduct of the accumulation of many small, independent changes along different parts of the genome and phenotype, without requiring any single dramatic mutation or reorganisation. And it provides empirical support for the fundamental premise of geographic (allopatric) speciation theory: that the reduction of gene flow by geographic distance or barriers is sufficient, given enough time, to produce new species.3, 15, 19
Ring species also highlight the inherent difficulty of applying a strict species concept to populations in the process of diverging. The terminal populations of a ring species are clearly distinct species by any criterion based on reproductive isolation, yet they are connected by a continuous chain of interbreeding populations through which genes could, in principle, flow from one end to the other. This creates a paradox for the biological species concept: two populations that do not interbreed (and are therefore separate species) are connected by gene flow through intermediates (and are therefore the same species). The resolution, as Mayr and subsequent authors have noted, is that the ring species simply represents an intermediate stage in the speciation process, one in which the gradual severing of gene flow is not yet complete at all points along the chain but has progressed far enough at the terminal overlap to produce functional species.2, 14, 15
The study of ring species has also contributed to the development of landscape genetics and phylogeography as disciplines. The analytical tools and conceptual frameworks developed to test ring species predictions — measuring genetic distance as a function of path distance around a barrier, quantifying clinal variation in multiple traits simultaneously, and using molecular phylogenetics to reconstruct the history of range expansion — have been applied broadly to the study of geographic variation and speciation in many other organisms.1, 17 Even in systems that are not ring species, the principle that geographic structure and barriers to gene flow shape the pattern and pace of divergence remains central to modern speciation research.3, 19
References
Phylogeography of the Ensatina eschscholtzii complex: parapatric intergradation and the ring species concept
Phylogeography of the herring gull complex (Larus argentatus) revealed by mitochondrial DNA and microsatellites
Genomic variation in a widespread Neotropical bird reveals divergence, population expansion, and gene flow
Clinal variation in the greenish warbler complex around the Tibetan Plateau: implications for speciation
Mitochondrial DNA phylogeography of the Ensatina eschscholtzii complex in coastal California