Overview
- Amino acid racemization dating exploits the slow, temperature-dependent conversion of L-amino acids to their D-enantiomers after the death of an organism, providing a chemical clock applicable to shells, bones, teeth, and other proteinaceous materials spanning the last few million years.
- Because the rate of racemization is strongly influenced by long-term temperature history, the technique functions as both a geochronometer and a paleothermometer — but this temperature sensitivity also means that independent calibration against radiometric or other absolute dates is essential for reliable age determination.
- When properly calibrated and cross-checked against methods like radiocarbon, uranium-series, and luminescence dating, amino acid racemization provides a cost-effective tool for correlating marine and terrestrial Quaternary sequences across wide geographic areas.
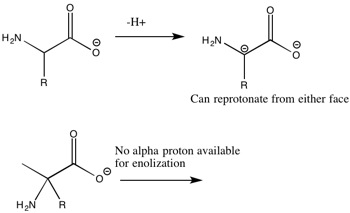
Amino acid racemization (AAR) is a chemical dating technique that exploits the slow, spontaneous interconversion of amino acid enantiomers in biological tissues after death. In living organisms, proteins are built almost exclusively from L-amino acids. Once an organism dies and its metabolic repair mechanisms cease, L-amino acids begin to convert to their D-enantiomeric forms through a process called racemization (or epimerization, in the case of amino acids like isoleucine that have two chiral centres). Over time, the ratio of D to L forms increases from near zero toward an equilibrium value of 1.0, and the extent of this conversion provides a measure of the time elapsed since death.1, 2 First applied to geochronology in the late 1960s by Philip Hare and Richard Mitterer and developed extensively by Jeffrey Bada through the 1970s and 1980s, the method has been applied to mollusc shells, foraminifera, coral, bones, teeth, and eggshells across the Quaternary and, in favourable cases, back several million years.1, 2, 4
{kind=link}
Chemical basis and kinetics
Racemization occurs because the alpha-carbon of an amino acid can undergo inversion of its stereochemical configuration through a reversible chemical reaction. The rate of this reaction differs among amino acids: aspartic acid racemizes relatively quickly, with detectable D/L ratios developing within centuries, while isoleucine epimerizes more slowly and is useful over longer timescales extending to several million years.2, 3 The kinetics of racemization are complex. Early workers modelled the reaction as a simple reversible first-order process, but subsequent research demonstrated that the effective rate depends on the molecular environment of the amino acid — whether it is in a free state, in a short peptide chain, or still bound within an intact protein — and that hydrolysis of the peptide bond and racemization are coupled reactions.2, 5
The most critical variable governing racemization rate is temperature. The reaction follows Arrhenius kinetics, with an activation energy of approximately 110–140 kJ/mol for most amino acids, meaning that a few degrees of long-term average temperature difference can produce large differences in D/L ratio for samples of the same age.2, 14 This temperature sensitivity is a double-edged feature of the method: it makes AAR useful as a paleothermometer when the age of a sample is independently known, but it also means that without independent temperature constraints, the method cannot yield absolute ages. For this reason, AAR is most commonly used as a relative dating and correlation tool (aminostratigraphy) or is calibrated against independent numerical ages from radiometric or luminescence dating.4, 6
The closed-system approach
A significant advance in AAR dating came with the recognition that amino acids trapped within the mineral crystal lattice of biominerals — the intra-crystalline fraction — behave as a closed system, isolated from the open exchange of amino acids with the surrounding environment that plagues analyses of the whole shell or bone.8, 9 Kirsty Penkman and colleagues demonstrated that by isolating this intra-crystalline protein decomposition (IcPD) fraction through bleaching the shell exterior with concentrated sodium hypochlorite, the resulting D/L ratios are far more reproducible and less susceptible to contamination or leaching than whole-shell measurements.8, 9 The closed-system behaviour can be tested empirically by subjecting samples to artificial heating experiments and verifying that amino acids are not lost from the bleached fraction, even at elevated temperatures that would cause massive open-system loss from unbleached material.9
The IcPD approach has revitalised aminostratigraphy as a practical correlation tool, particularly in the British Quaternary. Penkman and colleagues applied it to opercula of the freshwater gastropod Bithynia tentaculata, establishing an aminostratigraphy for UK interglacial deposits that discriminates marine isotope stages from MIS 1 through MIS 15 and beyond, independently confirming and extending the chronological framework provided by biostratigraphy and luminescence dating.10, 12 Similar approaches have been applied to Mediterranean marine terraces using Glycymeris and other mollusc genera, providing aminostratigraphic correlations of Quaternary shoreline deposits across thousands of kilometres of coastline.6, 11
Applications and materials
Mollusc shells remain the most widely used material for AAR dating, because their dense aragonite or calcite matrix preserves indigenous amino acids for hundreds of thousands to millions of years, and because molluscs are abundant in both marine and terrestrial Quaternary deposits.4, 7 John Wehmiller's extensive work on Atlantic and Pacific coastal raised beaches demonstrated that D/L ratios in molluscan genera such as Mercenaria and Saxidomus could be used to correlate terraces across the eastern and western seaboards of North America when calibrated against uranium-series coral ages from the same deposits.4, 7 Foraminifera from deep-sea sediment cores have also been dated by AAR, providing independent chronological checks on oxygen isotope stratigraphy in cores from the Atlantic, Pacific, and Indian Oceans.16
Bones and teeth present greater challenges because their porous, open microstructure allows amino acids to leach in and out over time, violating the closed-system assumption, and because microbial colonisation of bone can introduce exogenous amino acids that distort the indigenous racemization signal.5, 13 Nonetheless, careful selection of dense cortical bone or dentine, combined with the IcPD approach where applicable, has yielded useful results in archaeological and paleoanthropological contexts.2, 5 The extent of racemization in fossil bone has also proven useful as a screening criterion for ancient DNA preservation: Hendrik Poinar and colleagues showed that samples with D/L aspartic acid ratios above approximately 0.10 are unlikely to contain amplifiable endogenous DNA, providing a quick triage tool for ancient DNA studies before expensive extraction protocols are attempted.15
Calibration and concordance
Because racemization rates are temperature-dependent and non-linear, AAR cannot function as a stand-alone absolute dating method without calibration. The standard approach is to measure D/L ratios in samples of independently known age from the same region, establishing a local calibration curve that implicitly accounts for the integrated thermal history of that area.2, 4 Radiocarbon dating calibrates the youngest part of this curve (to approximately 50,000 years), while uranium-series dating of corals and luminescence dating of associated sediments extend the calibration through the Middle and Late Pleistocene.6, 7 Once calibrated, the D/L ratio of an unknown sample from the same geographic and thermal regime can be interpolated to yield a numerical age estimate, typically with uncertainties of 10–20 percent.4
The concordance of AAR ages with those from independent methods is the ultimate test of the technique's validity. In the Mediterranean and on the Atlantic coastal plain of North America, aminostratigraphic assignments based on D/L ratios consistently agree with uranium-series, luminescence, and electron spin resonance ages on the same or correlative deposits, confirming that the kinetic models and calibrations are sound within their applicable temperature and time ranges.6, 7, 10 Where discrepancies arise, they have typically been traced to open-system behaviour in poorly preserved samples or to inadequate accounting for thermal history differences between calibration and unknown sites — problems that the closed-system IcPD approach has largely resolved for molluscan materials.8, 9 As both a relative correlation tool and, when properly calibrated, a numerical dating method, amino acid racemization continues to fill a valuable niche in the geochronological toolkit, complementing the radiometric and physical methods that dominate Quaternary chronology.2, 10
References
Kinetics of amino acid racemization (epimerization) in the dentine of fossil and modern bear teeth
Aminostratigraphy and geochronology of Quaternary shorelines in the Mediterranean basin
Amino acid racemization dating of late Pleistocene mollusc shells from the Palos Verdes Hills, California
Intra-crystalline protein decomposition (IcPD): a new dating method for Quaternary Mollusca
Closed-system behaviour of the intra-crystalline fraction of amino acids in mollusc shells
Dating of raised marine terraces in the eastern Mediterranean by amino acid racemization
Amino acid geochronology: a closed-system approach to test and refine the UK aminostratigraphy
Racemization of isoleucine in fossil molluscs from Indian raised beaches and implications for uplift rate
Temperature and water activity as major controls on aspartic acid racemization in biological tissues