Overview
- Anatomically modern humans evolved in Africa by at least 300,000 years ago and made several early forays into the Levant and Arabia before a major dispersal wave between roughly 70,000 and 50,000 years ago seeded all non-African populations, producing a characteristic gradient of declining genetic diversity with increasing distance from the continent.
- The dispersal involved multiple routes and episodes of interbreeding with archaic hominins: all non-African populations carry approximately 1 to 4 percent Neanderthal DNA from contact in western Asia, while Melanesian and Aboriginal Australian populations carry an additional 3 to 6 percent Denisovan ancestry acquired in Southeast Asia or Near Oceania.
- Humans reached Australia by roughly 65,000 years ago, Europe by 46,000 years ago, the Americas by at least 16,000 years ago, and Remote Oceania within the past 3,500 years, with Pacific island colonization representing the final chapter of a global dispersal accomplished entirely during the Pleistocene and Holocene.
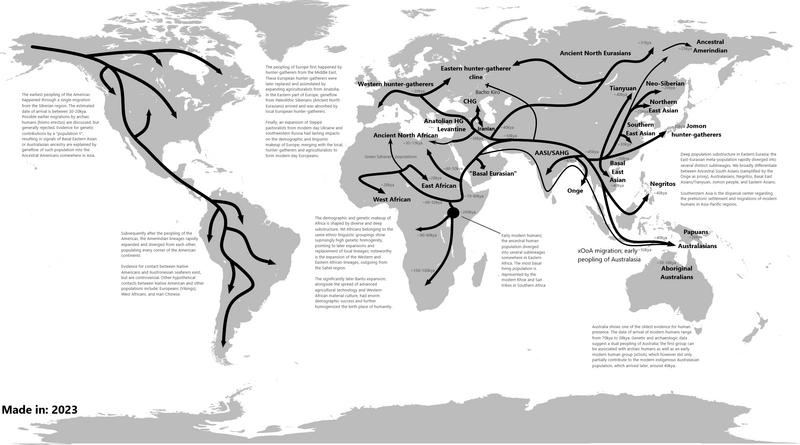
Modern human dispersal refers to the process by which Homo sapiens, having evolved in Africa by at least 300,000 years ago, spread across the entire habitable world over the course of roughly 100,000 years.2 This expansion was not a single, coordinated migration but a complex series of population movements, dead-end forays, bottlenecks, and secondary expansions shaped by climate, geography, and encounters with archaic hominin populations already resident in Eurasia. The genetic, fossil, and archaeological evidence now available makes it possible to reconstruct the dispersal in considerable detail, from the earliest tentative exits into the Levant to the colonization of Remote Oceania within the past few millennia.1, 6 Understanding this dispersal is central to explaining the pattern of ancient DNA and population history observed among living humans: the gradient of declining genetic diversity with increasing distance from Africa, the distribution of archaic ancestry, and the deep population structure that underlies regional differences.6
African origins and the source population
The dispersal of modern humans presupposes an origin. Fossil and genetic evidence converge on the conclusion that Homo sapiens evolved in Africa, with the oldest known fossils attributed to our species coming from Jebel Irhoud in Morocco (approximately 315,000 years ago) and the Omo Kibish Formation in Ethiopia (approximately 233,000 years ago).2 Rather than arising in a single geographic cradle, early Homo sapiens in Africa appear to have evolved across a continent-wide network of subdivided populations connected by intermittent gene flow, a process sometimes called African multiregionalism. The anatomically modern suite of features — a globular braincase, retracted face, and prominent chin — assembled gradually in a mosaic fashion over more than 200,000 years.2
The source population for the out-of-Africa dispersal was a subset of this broader African diversity. Genetic analyses consistently show that the deepest branches of the human phylogenetic tree separate African populations from one another — particularly the Khoisan-speaking peoples of southern Africa and certain East African groups — while all non-African populations cluster together on a comparatively shallow branch.1, 6 This topology indicates that non-Africans derive from a population that split from other African lineages relatively recently, between roughly 70,000 and 50,000 years ago, and then expanded rapidly with a dramatically reduced effective population size. The serial founder effect produced by successive small groups budding off from the expanding wavefront explains the smooth, predictable decline in heterozygosity observed from East Africa to the tip of South America.6
Early dispersals into the Levant and Arabia
The main out-of-Africa dispersal was preceded by multiple earlier forays that left little or no lasting genetic legacy in present-day populations. The oldest widely cited evidence comes from a partial cranium at Apidima Cave in southern Greece, tentatively attributed to Homo sapiens and dated to approximately 210,000 years ago, though both the identification and the dating remain contested.17 More securely established is the evidence from the Levant. A maxilla from Misliya Cave in Israel, dated to approximately 177,000–194,000 years ago, provides the earliest broadly accepted fossil evidence of modern humans outside Africa.3
The Skhul and Qafzeh cave sites in Israel have yielded a substantial assemblage of anatomically modern human fossils dated to approximately 90,000–120,000 years ago. These populations occupied the Levant during a warm interglacial interval (marine isotope stage 5), apparently overlapping temporally with Neanderthals in the broader region. The Skhul/Qafzeh humans used Mousterian stone tool technology similar to that of contemporary Neanderthals, and they appear to have been displaced or to have died out by around 70,000 years ago, when Neanderthals reoccupied the Levantine caves.4 This pattern suggests that the early Levantine presence of Homo sapiens represented a climate-dependent range expansion rather than a permanent colonization.
The Arabian Peninsula provides additional evidence of early dispersals. A Homo sapiens finger bone from the site of Al Wusta in the Nefud Desert of Saudi Arabia, dated to approximately 85,000 years ago, demonstrates that modern humans penetrated deep into the Arabian interior during a humid phase when the peninsula supported lakes, rivers, and grasslands.5 Stone tool assemblages across the Arabian Peninsula and the Horn of Africa suggest that these early dispersals tracked green corridors that opened periodically across the Sahara and into the Levant and Arabia during favorable climatic intervals. Most of these early populations appear to have left no detectable descendants among living non-African humans, though some genomic studies have identified faint traces of a pre-main-dispersal signal in certain Australasian genomes.18, 19
The main out-of-Africa expansion
The dispersal event responsible for founding all major non-African populations occurred between approximately 70,000 and 50,000 years ago. Genomic analyses of living and ancient populations consistently point to a severe population bottleneck during this period, when the effective population size of the emigrating group may have dropped to only a few thousand individuals before expanding rapidly across southern Asia, Southeast Asia, and eventually into Europe and the Americas.6, 8 The bottleneck is written into every non-African genome: compared to African populations, non-Africans display markedly lower heterozygosity, longer haplotype blocks (reflecting fewer generations of recombination since the founding event), and an excess of recently derived, population-specific mutations consistent with rapid post-bottleneck expansion.6
.png){kind=link}
Two primary routes out of Africa have been proposed. The northern route follows the Sinai Peninsula through the Levant into western Asia, while the southern route crosses the Bab el-Mandeb strait at the mouth of the Red Sea into the Arabian Peninsula and proceeds along the Indian Ocean coastline toward South and Southeast Asia. The southern coastal route gained popularity because it provides a mechanism for the remarkably early colonization of Australia by approximately 65,000 years ago, implying rapid, directed movement along a coastal corridor.7, 18 However, the archaeological record of this proposed coastal migration is almost entirely missing, in part because Pleistocene shorelines are now submerged under 60 to 120 metres of water.
Genomic evidence suggests that the distinction between northern and southern routes may be an oversimplification. A 2016 analysis of 483 diverse human genomes found that most non-African populations derive from a single major dispersal wave, with Papuan genomes carrying a small additional ancestry component (~2%) from an earlier, separate dispersal.19 The Aboriginal Australian genome study of the same year demonstrated that the ancestors of Aboriginal Australians diverged from the ancestors of mainland Eurasians approximately 58,000 years ago, consistent with a single dispersal that subsequently split into geographically distinct branches rather than two wholly independent migrations.8 The current consensus favors a predominantly single dispersal with possible minor contributions from earlier exits detectable in certain Oceanian populations.
Colonization of South Asia, Southeast Asia, and Australia
The route from western Asia to Australia required traversing the full breadth of southern Asia and navigating the island archipelagos of Southeast Asia, including water crossings that no hominin had previously accomplished. The speed of this expansion was remarkable: if modern humans left Africa around 65,000–70,000 years ago and reached northern Australia by approximately 65,000 years ago, as indicated by the Madjedbebe rock shelter in the Northern Territory, the crossing of more than 15,000 kilometres of coastline and open water was accomplished in at most a few thousand years.7
The colonization of the Australian continent (then connected to New Guinea as the landmass known as Sahul) required multiple sea crossings through the islands of Wallacea, where even at times of maximum sea-level lowering during glacial periods, individual crossings of at least 60 to 90 kilometres of open water were necessary. The ability to make these crossings implies watercraft technology, though no direct archaeological evidence of Pleistocene boats has survived.7 Genomic analyses of Aboriginal Australian populations reveal deep internal divergence dating to approximately 50,000 years ago, indicating that once Sahul was colonized, populations dispersed rapidly across the continent and subsequently remained relatively isolated from later Eurasian population movements for tens of thousands of years.8 A later Holocene gene flow event from the Indian subcontinent, dated to approximately 4,000 years ago, has been detected in some Aboriginal Australian genomes, coinciding with the appearance of the dingo and changes in stone tool technology.24
An early genomic study of a hair sample from a Western Australian Aboriginal individual demonstrated that Aboriginal Australians descend from an early dispersal wave into Asia and are not derived from later East Asian or European populations, supporting the antiquity and distinctness of the Sahul founding population.20 The presence of both Neanderthal and Denisovan ancestry in Aboriginal Australian and Melanesian genomes indicates that the ancestors of these populations encountered and interbred with archaic hominins during their transit through mainland and island Southeast Asia.8, 11
European colonization
Europe was colonized later than Australia, despite its closer proximity to Africa. Modern humans reached southeastern Europe by approximately 46,000–45,000 years ago, as documented by Homo sapiens fossils and Initial Upper Palaeolithic (IUP) artifacts from Bacho Kiro Cave in Bulgaria.13 The IUP toolkit, characterized by elongated blade technologies with some retention of Middle Palaeolithic elements, represents the earliest archaeological signal of Homo sapiens in Europe and appears at sites from the Levant through southeastern Europe.

{kind=link}
The colonization was a protracted process rather than a rapid sweep. Modern humans initially entered through the Mediterranean and Danubian corridors, gradually expanding northward and westward. For several thousand years, Homo sapiens and Neanderthals occupied different parts of Europe, with Neanderthals persisting in refugia on the Iberian Peninsula and in parts of western Europe until approximately 40,000 years ago.10, 13 Genetic evidence from early European Homo sapiens specimens reveals that many of these earliest colonizers left no detectable descendants among later Europeans. The Oase individual from Romania, dated to approximately 40,000 years ago, carried 6 to 9 percent Neanderthal DNA — indicating a Neanderthal ancestor within the preceding four to six generations — but does not appear to have contributed ancestry to any later European population.10
By approximately 35,000 years ago, the Upper Palaeolithic was fully established across Europe, associated exclusively with Homo sapiens. The Aurignacian, Gravettian, and subsequent technocomplexes are distinguished by prismatic blade production, bone and antler tools, figurative art, musical instruments, and elaborate personal ornaments — a cultural florescence that has no parallel in the preceding Neanderthal-associated Mousterian record.13 The Last Glacial Maximum (approximately 26,000–19,000 years ago) caused significant population contraction, with surviving groups retreating to southern refugia in Iberia, Italy, and the Balkans before re-expanding northward as conditions ameliorated.
Interbreeding with Neanderthals and Denisovans
The dispersal of modern humans brought them into contact with archaic hominin populations that had occupied Eurasia for hundreds of thousands of years. Genomic analyses have demonstrated unequivocally that interbreeding occurred, and the resulting archaic introgression has left a measurable signature in the genomes of all living non-African populations.9
The sequencing of the Neanderthal genome revealed that non-African humans carry approximately 1 to 4 percent Neanderthal-derived DNA, while sub-Saharan Africans carry essentially none, placing the interbreeding event after the out-of-Africa dispersal but before the differentiation of major non-African populations.9 The most probable location and timing for this admixture is western Asia between approximately 60,000 and 50,000 years ago, when expanding Homo sapiens first encountered resident Neanderthal groups in a zone of overlap. The consistency of Neanderthal ancestry proportions across geographically diverse non-African populations — from Europeans to East Asians to Native Americans — indicates that the admixture occurred before these populations diverged from one another.9, 21
Denisovan admixture follows a different geographic pattern. The highest levels of Denisovan ancestry, approximately 3 to 6 percent, are found in Melanesian, Aboriginal Australian, and some Philippine Negrito populations, while mainland East Asian populations carry substantially less (on the order of 0.2 percent).11, 21 Genomic analyses of Papuan populations have revealed at least two distinct episodes of Denisovan admixture involving genetically differentiated Denisovan populations, suggesting that Homo sapiens encountered Denisovans at multiple locations or times during the transit through Southeast Asia and into Oceania.12 Many introgressed archaic alleles have been retained by natural selection because they conferred adaptive advantages in the environments modern humans were entering, including Neanderthal-derived immune genes and the Denisovan-derived EPAS1 allele that facilitates high-altitude adaptation in Tibetan populations.9, 21
Peopling of the Americas
The Americas were the last major continental landmass to be colonized by Homo sapiens. Entry required crossing Beringia, the land bridge connecting northeastern Asia to Alaska that was exposed during glacial periods of lowered sea level. Genetic and archaeological evidence converges on an entry date of at least 16,000 years ago, though the recent discovery of human footprints at White Sands, New Mexico, dated to approximately 23,000 years ago, has raised the possibility of a substantially earlier presence that remains under active debate.14, 23
The Beringian standstill model, proposed on the basis of mitochondrial DNA and later supported by whole-genome analyses, holds that the ancestors of Native Americans diverged from East Asian populations approximately 25,000 to 20,000 years ago and spent several thousand years genetically isolated in Beringia before expanding south into the Americas proper.15 This period of isolation allowed the accumulation of private mutations that distinguish Native American lineages from their closest Asian relatives. The route of southward expansion has been debated: the traditional model posited an ice-free corridor between the Laurentide and Cordilleran ice sheets, but the chronology and ecology of this corridor make it unlikely to have been viable before about 13,000 years ago. The alternative Pacific coastal route, following a "kelp highway" of productive marine habitats along the western coast, better explains the early appearance of sites like Monte Verde in southern Chile, securely dated to approximately 14,500 years ago.14, 16
Ancient DNA from early American remains has revealed that all Indigenous peoples of the Americas south of the Arctic descend primarily from a single founding population, which subsequently diversified into northern (Ancestral A) and southern (Ancestral B) branches. At least two additional migration waves from Asia contributed ancestry to specific populations: one associated with Na-Dene speakers and another with Inuit and other Arctic peoples.14 The speed of the initial colonization was extraordinary: within approximately 1,500 years of entering the continent, humans had reached the southern tip of South America, more than 14,000 kilometres from the point of entry.16
Island colonization and the Pacific
The colonization of the Pacific islands represents the final major chapter of human dispersal and one of the most remarkable feats of long-distance navigation in prehistory. Near Oceania — encompassing New Guinea, the Bismarck Archipelago, and the Solomon Islands — was settled during the initial Pleistocene expansion, with archaeological evidence of human occupation dating to approximately 45,000 years ago in New Britain and New Ireland. These crossings, though shorter than those required for reaching Australia, still involved navigation across open water gaps of 30 to 90 kilometres.22
Remote Oceania — the vast expanse of the Pacific beyond the Solomon Islands, including Vanuatu, Fiji, Tonga, Samoa, and eventually Polynesia — remained uninhabited until the expansion of Austronesian-speaking peoples beginning approximately 3,300 to 3,000 years ago. This expansion is associated archaeologically with the Lapita cultural complex, distinguished by its distinctive dentate-stamped pottery, and genetically with a population that originated in Taiwan and moved through the Philippines and into Melanesia before striking out into the open Pacific.22, 26 Ancient DNA from early Lapita burials in Vanuatu and Tonga has revealed that the first settlers of Remote Oceania had predominantly East Asian ancestry with minimal Papuan admixture, though subsequent gene flow from Near Oceanian populations increased the Papuan ancestry component substantially in many island groups over the following centuries.26
The colonization of the most remote Pacific islands occurred in successive waves. Eastern Polynesia — including the Marquesas, Society Islands, and Hawai'i — was settled between approximately 1,000 and 800 years ago. New Zealand, the largest and most southerly landmass colonized by Polynesian voyagers, was reached approximately 700 years ago. These voyages were accomplished using double-hulled canoes navigated by stellar, wave, and current observations, covering distances of thousands of kilometres across open ocean.25 The colonization of Remote Oceania closed the final habitable gap on the globe: by the time European maritime exploration began in the fifteenth century, Homo sapiens had already reached virtually every inhabitable island in the Pacific.
Genetic signatures of the dispersal
The global dispersal of Homo sapiens is recorded in the genomes of living and ancient populations through several distinct genetic signatures. The most fundamental is the serial founder effect: as small groups budded off from the expanding wavefront and colonized new territory, each successive founding population carried only a fraction of the genetic diversity present in the source population. This process produced a remarkably smooth, predictable decline in heterozygosity with increasing geographic distance from East Africa, a pattern confirmed by studies of autosomal DNA, mitochondrial DNA, Y-chromosome lineages, and even phenotypic traits such as cranial shape diversity.6
Mitochondrial DNA and Y-chromosome phylogenies provide complementary perspectives on the dispersal. All non-African mitochondrial lineages belong to the macro-haplogroups M and N, both of which descend from the L3 haplogroup that originated in Africa. The coalescence dates of M and N, estimated at approximately 60,000–70,000 years ago, provide a molecular clock estimate for the out-of-Africa bottleneck that is broadly consistent with the archaeological and fossil record.1 Y-chromosome phylogenies tell a similar story, with all non-African male lineages descending from a subset of African haplogroups that diversified after the dispersal.
Whole-genome analyses have added resolution to these patterns. Haplotype block lengths are longer outside Africa, reflecting fewer generations of recombination since the founding bottleneck. The site frequency spectrum of non-African populations shows an excess of rare, recently derived variants, consistent with rapid demographic expansion from a small founding group. Runs of homozygosity are longer and more frequent in populations at greater geographic distances from Africa, a direct consequence of the serial reduction in effective population size with each successive founding event.6, 8 These genome-wide patterns are so regular and predictable that geographic distance from East Africa alone explains the majority of variance in genetic diversity among human populations, a finding that underscores the dominant role of the dispersal process in shaping modern human genetic architecture.
Synthesis
The dispersal of Homo sapiens from Africa to every habitable corner of the globe was accomplished in a geologically brief span of time. From the earliest tentative forays into the Levant more than 100,000 years ago, through the main expansion between 70,000 and 50,000 years ago, to the colonization of Remote Oceania within the past millennium, the process involved repeated episodes of population movement, bottleneck, expansion, and admixture with both archaic hominins and previously established modern human groups.6, 9 The genetic consequences of this dispersal — the gradient of declining diversity, the distribution of archaic ancestry, the signatures of founder effects and admixture — remain clearly legible in the genomes of living people and provide the foundation for understanding human population history, adaptation, and diversity.1, 6, 8
References
The dispersal of Homo sapiens across southern Asia: how early, how often, how complex?
Footprints in White Sands National Park provide evidence for human presence in the Americas circa 23,000 years ago