Overview
- Diplodocus was a massive diplodocid sauropod dinosaur of the Late Jurassic Morrison Formation, reaching body lengths of 24 to 26 metres with a distinctive whip-like tail and pencil-shaped teeth adapted for a specialized stripping or raking feeding strategy.
- The Carnegie Museum’s mounted skeleton of Diplodocus carnegii, known as ‘Dippy,’ became the most widely exhibited dinosaur in history when plaster casts were donated to major museums across the world in the early twentieth century, making it the first dinosaur most people ever saw.
- The former genus Seismosaurus, once claimed as the longest dinosaur ever discovered at over 50 metres, was reclassified as a species of Diplodocus (D. hallorum), with revised length estimates of approximately 32 to 33 metres — still enormous, but far shorter than the original sensationalized claims.
Diplodocus is a genus of diplodocid sauropod dinosaur that lived during the Late Jurassic period, approximately 154 to 150 million years ago, in what is now western North America. First described by Othniel Charles Marsh in 1878 from vertebrae recovered from the Morrison Formation of Colorado, Diplodocus is one of the most completely known sauropod dinosaurs and among the most iconic prehistoric animals in the world, owing largely to the famous Carnegie Museum skeleton whose plaster casts were donated to major museums across Europe, South America, and beyond in the early twentieth century.1, 2 With a total body length of 24 to 26 metres in the best-known species, D. carnegii, and estimated body masses of 11,000 to 16,000 kilograms depending on the modelling approach used, Diplodocus was one of the longest terrestrial animals for which complete skeletal material is available, though not the heaviest — a distinction that belongs to the more robustly built macronarian sauropods such as Brachiosaurus and the titanosaurs.14, 15
Carnegie Museum, Wikimedia Commons, No restrictions
_-_its_osteology,_taxonomy,_and_probable_habits,_with_a_restoration_of_the_skeleton_(1901)_(20964449321).jpg){kind=link}
The genus is characterised by an extraordinarily elongate body plan dominated by a very long neck and an even longer, whip-like tail, supported by a relatively compact torso and columnar limbs. The teeth of Diplodocus are pencil-shaped and confined to the anterior portion of the jaws, a specialised dental morphology that indicates a feeding strategy fundamentally different from that of the spoon-toothed sauropods like Camarasaurus with which it coexisted. Three valid species are currently recognised by most workers: D. longus (the type species), D. carnegii, and D. hallorum (formerly Seismosaurus hallorum), though the taxonomy and species-level systematics of the genus remain subjects of active investigation.14, 16
Discovery and naming
The first fossils of Diplodocus were discovered by Samuel Wendell Williston and Benjamin Franklin Mudge in 1877 during expeditions sponsored by Othniel Charles Marsh of Yale College. The specimen, consisting of a series of caudal vertebrae and chevrons, was recovered from Morrison Formation exposures near Cañon City, Colorado. Marsh described and named the genus in 1878, erecting the type species Diplodocus longus ("long double beam") in reference to the distinctive double-beamed chevron bones on the underside of the tail vertebrae, which were unique among dinosaurs known at the time. The chevrons possess both anterior and posterior projections that together form a V-shaped or "double beam" structure, and it was this unusual morphology, rather than the animal's great length, that inspired the generic name.1
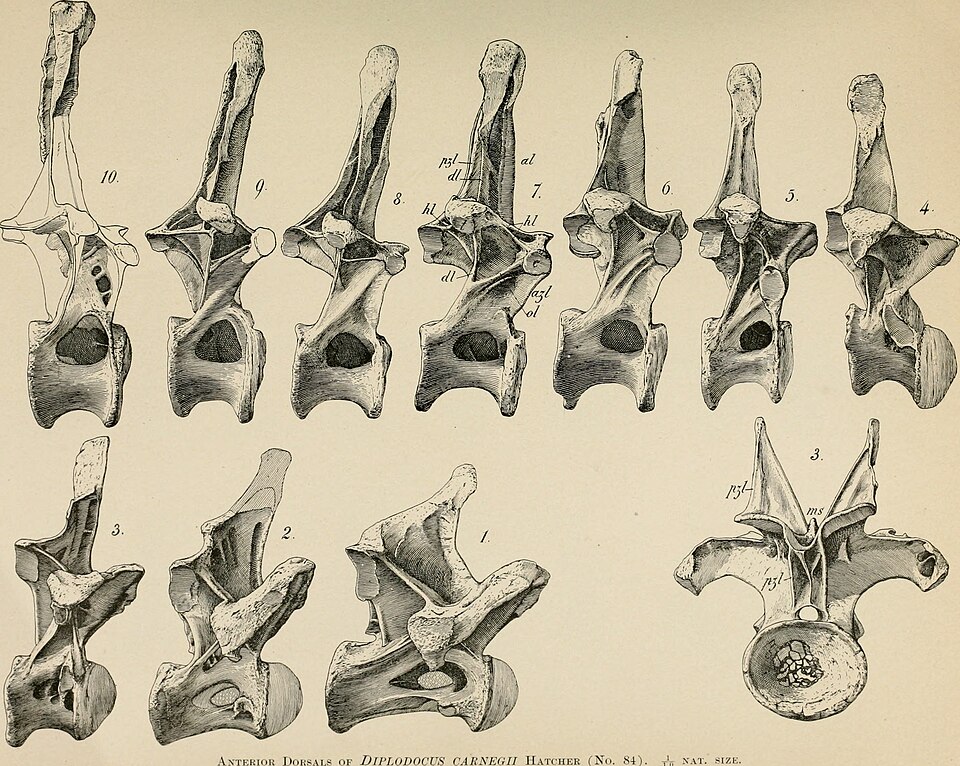
The far more complete and influential species D. carnegii was described by Hatcher in 1901 on the basis of a nearly complete skeleton (CM 84) discovered in 1899 at Sheep Creek, Albany County, Wyoming. The specimen was found by a team working under the sponsorship of Andrew Carnegie, the industrialist and philanthropist, who had become fascinated by newspaper reports of giant dinosaurs discovered in the American West. Carnegie funded extensive fieldwork in the Morrison Formation, and the resulting skeleton was mounted at the Carnegie Museum of Natural History in Pittsburgh, where it became one of the first sauropod skeletons to be displayed in its entirety.2
Carnegie subsequently commissioned plaster casts of the mounted skeleton and donated them to museums around the world, including the Natural History Museum in London (1905), the Muséum national d'Histoire naturelle in Paris (1908), the Naturkundemuseum in Berlin (1908), the Museo de La Plata in Argentina (1912), and several others. These casts, collectively known as "Dippy," introduced Diplodocus to millions of museum visitors on multiple continents and made it the first dinosaur that many people ever encountered. The London cast became so iconic that it remained the centrepiece of the Natural History Museum's main hall for over a century until its replacement in 2017, and it continues to tour British museums as a cultural ambassador for palaeontology.2, 3
Species and the Seismosaurus debate
The species-level taxonomy of Diplodocus has undergone substantial revision over the past two decades, with the most significant change being the reclassification of Seismosaurus hallorum as Diplodocus hallorum. Seismosaurus ("earth shaker lizard") was described in 1991 by David Gillette from a partial postcranial skeleton (NMMNH P-3690) recovered from the Morrison Formation of New Mexico. Gillette initially estimated the animal's total length at 39 to 52 metres, which would have made it the longest dinosaur ever discovered by a wide margin, and the sensational size estimates attracted enormous media attention and public interest.4
Subsequent study of the specimen revealed that the original length estimates were substantially inflated. Lucas and colleagues' 2006 reevaluation demonstrated that many of the vertebrae had been misidentified as to their position in the vertebral column, leading to an overestimate of the number of vertebrae and hence the length of the neck and tail. When the vertebrae were correctly reidentified and the skeleton was reconstructed using more accurate proportional data from other diplodocids, the revised length estimate fell to approximately 32 to 33 metres — still very large, but well within the size range of other long diplodocids such as Supersaurus. Furthermore, Lucas and colleagues concluded that the anatomical features that distinguished Seismosaurus from Diplodocus fell within the range of individual variation expected within a single genus, and they formally reclassified the taxon as Diplodocus hallorum.4
Tschopp, Mateus, and Benson's 2015 comprehensive specimen-level phylogenetic analysis of Diplodocidae, which scored 477 characters across 81 operational taxonomic units, supported the inclusion of D. hallorum within Diplodocus while also recovering D. longus and D. carnegii as valid species. The analysis placed Diplodocus within the subfamily Diplodocinae alongside Barosaurus, Kaatedocus, and Leinkupal, with Apatosaurinae (Apatosaurus, Brontosaurus) recovered as the sister group. This study also controversially resurrected the genus Brontosaurus as distinct from Apatosaurus, demonstrating that specimen-level phylogenetic analysis can resolve long-standing taxonomic questions that traditional approaches could not.16
Anatomy and the whip tail
The overall body plan of Diplodocus is dominated by extreme axial elongation, with the neck and tail together accounting for approximately 80 percent of the animal's total length. The neck of D. carnegii contains 15 elongate cervical vertebrae, each deeply excavated by pneumatic fossae (air-filled cavities) that reduced the mass of the vertebrae while maintaining their structural integrity through a system of thin bony laminae. This pneumatic construction is a hallmark of sauropod vertebral architecture and is homologous with the postcranial pneumaticity observed in modern birds, representing air sacs that extended from the respiratory system into the skeleton.12, 18
The tail of Diplodocus is among the longest of any known dinosaur, comprising approximately 80 caudal vertebrae in D. carnegii. The proximal (base) caudal vertebrae are large and bear the characteristic double-beamed chevrons that inspired the generic name, but the vertebrae become progressively smaller toward the tip, with the distal third of the tail consisting of tiny, rod-like vertebrae only a few centimetres in length. This tapering structure gives the tail a whip-like profile, and Myhrvold and Currie's 1997 biomechanical analysis demonstrated that a diplodocid tail of these proportions could, if swung laterally with the base held relatively still, achieve velocities at the tip exceeding the speed of sound (approximately 340 metres per second), producing a supersonic crack analogous to the snap of a bullwhip.7
The functional significance of a supersonic tail tip remains debated. Myhrvold and Currie suggested that the sound produced by a supersonic tail snap could have served as an acoustic signal — for species recognition, mate attraction, or predator deterrence — or that the tail could have been used as a defensive weapon against predators such as Allosaurus. Sceptics have noted that the tiny, fragile distal vertebrae show no obvious adaptations for withstanding the extreme forces generated by supersonic motion, and that the energetic cost of achieving such velocities may have been prohibitive. Nevertheless, the whip-like tail of Diplodocus remains one of the most distinctive and biomechanically intriguing features of any sauropod.7
The skull of Diplodocus is small relative to body size, measuring approximately 60 centimetres in length, with a gently sloping profile and nares (nasal openings) positioned high on the skull near the top of the head. The teeth are pencil-shaped (cylindrical with chisel-like tips), confined to the anterior portion of the premaxilla and dentary, and oriented nearly horizontally rather than vertically. This dental morphology is radically different from the broad, spoon-shaped teeth of macronarian sauropods such as Camarasaurus and Brachiosaurus, and it indicates a specialised feeding mechanism in which vegetation was stripped from branches by a raking or combing motion of the teeth rather than being bitten off or cropped.9, 11
The neck posture debate
The habitual posture of the neck in Diplodocus and other sauropods has been one of the most contentious issues in sauropod palaeobiology. Two fundamentally opposed positions have been advanced: the "horizontal neck" model, in which the neck was held roughly level with the back or angled slightly downward, and the "raised neck" model, in which the neck was held in an elevated, swan-like posture that would have given the animal access to high-browse vegetation.
Barrett and Upchurch's 2005 analysis of the osteological neutral pose (ONP) of sauropod necks — the posture in which the cervical vertebrae articulate with minimum cartilage interposition — found that the ONP of Diplodocus places the neck in a roughly horizontal to slightly downward-angled posture, with the head positioned at approximately the level of the shoulders or lower. This result supported the view that Diplodocus was primarily a low-browser, feeding at or below shoulder height by sweeping its long neck in broad arcs to cover a large feeding envelope without moving its massive body.5
Taylor, Wedel, and Naish's 2009 study challenged this interpretation by examining neck posture in extant animals. They demonstrated that virtually all living terrestrial vertebrates — mammals, birds, and reptiles — habitually hold their necks well above the ONP, with the head elevated substantially above the level of the back. If this pattern is universal among living amniotes, they argued, then there is no reason to assume that sauropods were exceptions, and the ONP of the vertebrae does not reliably predict the habitual posture of the living animal. They concluded that Diplodocus most likely held its neck in a raised posture at least some of the time, giving it access to browse at heights of 5 to 6 metres or more, though the neck was probably not held in a near-vertical position like that of Brachiosaurus.6
The debate remains unresolved and is complicated by the difficulty of reconstructing soft tissue anatomy — particularly the intervertebral cartilage, ligaments, and musculature — in an animal for which no close living analogues exist. The compromise view held by many workers is that Diplodocus was capable of both raised and horizontal neck postures, employing different postures for different activities: a raised posture for high browsing when profitable, and a horizontal or lowered posture for ground-level feeding or drinking. The extreme length of the neck would have provided flexibility in feeding strategy regardless of the habitual resting posture.5, 6, 18 Taylor and Wedel's 2013 analysis of why sauropods evolved long necks argued that the primary advantage was energetic: a long neck allowed the animal to feed over a large area without moving its enormous body, reducing the metabolic cost of locomotion. This "sessile feeding" strategy would have been particularly advantageous for Diplodocus, whose relatively low body mass for its length made it efficient at sweeping a broad feeding envelope while remaining stationary, in contrast to the higher, more vertically oriented necks of macronarians that prioritised access to tall vegetation.18
Feeding ecology
The pencil-shaped teeth and anterior-restricted dentition of Diplodocus indicate a feeding mechanism fundamentally different from that of the broad-toothed macronarian sauropods with which it coexisted. Upchurch and Barrett's comparative analysis of sauropod feeding mechanisms proposed that Diplodocus and other diplodocids employed a "branch stripping" or "unilateral branch stripping" technique, in which the animal clamped its teeth around a branch and pulled its head back, stripping leaves and small twigs from the stem. This interpretation is supported by the unusual wear patterns on diplodocid teeth, which show wear facets on only one side of the tooth crown (the labial side of the upper teeth and the lingual side of the lower teeth), consistent with a stripping motion rather than bilateral occlusion.11
The dietary niche of Diplodocus in the Morrison Formation ecosystem has been interpreted in the context of niche partitioning among the extraordinary diversity of sauropods present in the formation. At least five genera of large sauropods — Diplodocus, Apatosaurus, Barosaurus, Camarasaurus, and Brachiosaurus — coexisted in the Morrison Formation, representing a level of large-herbivore diversity unmatched in any modern ecosystem. The coexistence of so many ecologically similar species has been explained by niche partitioning along two primary axes: feeding height (with Brachiosaurus as a high-browser, Diplodocus and Apatosaurus as low to mid-level feeders, and Camarasaurus as a generalist) and food type (with the pencil-toothed diplodocids specialising on softer vegetation and the spoon-toothed Camarasaurus processing tougher plant material).5, 11, 14
Dental and feeding comparisons among Morrison Formation sauropods9, 11
| Genus | Tooth shape | Tooth position | Inferred feeding |
|---|---|---|---|
| Diplodocus | Pencil-shaped | Anterior only | Branch stripping |
| Apatosaurus | Pencil-shaped | Anterior only | Branch stripping |
| Camarasaurus | Spatulate (spoon) | Along jaw | Cropping, tougher plants |
| Brachiosaurus | Spatulate (spoon) | Along jaw | High browsing |
Reproduction and social behaviour
Direct evidence for the reproductive biology and social behaviour of Diplodocus is sparse, as is typical for sauropod dinosaurs, but indirect inferences can be drawn from nesting sites, trackways, and comparisons with other sauropods for which better data are available. No nesting sites have been definitively attributed to Diplodocus, but sauropod nesting grounds from other formations and continents — notably the titanosaur nesting sites of Auca Mahuevo in Argentina — indicate that sauropods laid clutches of relatively small eggs (typically 15 to 20 centimetres in diameter) in shallow excavations, buried them, and provided no parental care. If Diplodocus followed this pattern, hatchlings would have been extremely small relative to adult body size, perhaps 1 to 2 kilograms at hatching compared to 11,000 to 16,000 kilograms as adults — a growth factor of approximately 10,000-fold, among the highest known for any vertebrate.12, 14
Trackway evidence from the Morrison Formation provides some information about locomotion and potential group behaviour. Sauropod trackways from several Morrison Formation sites include parallel sets of tracks suggesting that multiple individuals were travelling together, though whether these represent coordinated herding behaviour or simply animals following the same trail to a shared resource (water, feeding area) cannot be determined from tracks alone. The absence of small tracks alongside large ones at most sites has been interpreted as evidence that juveniles and adults occupied different habitats or moved in separate groups, a pattern consistent with the hypothesis of ontogenetic niche partitioning proposed for other sauropod taxa.14, 16
The extreme size disparity between hatchling and adult Diplodocus implies that individuals passed through a wide range of ecological niches during their lifetime. A one-kilogram hatchling would have been vulnerable to predation by even small theropods and would have fed on entirely different vegetation than a 15,000-kilogram adult. This ontogenetic niche shift — in which juveniles and adults effectively function as different ecological species — is a distinctive feature of sauropod biology and has been proposed as one of the factors that limited sauropod species diversity in any given ecosystem, since each species effectively occupied multiple ecological roles across its lifetime.12
Growth rates and the Morrison Formation
The growth dynamics of Diplodocus have been investigated through bone histology, contributing to the broader understanding of how sauropod dinosaurs achieved and sustained their enormous body sizes. Curry's 1999 study of long bone histology in Diplodocus and Apatosaurus identified rapidly deposited fibrolamellar bone tissue in the cortex of juvenile and subadult individuals, indicating growth rates comparable to those of large modern mammals and far exceeding those of living reptiles. Lines of arrested growth (LAGs) in the cortex suggest that growth was seasonal and that skeletal maturity was reached at approximately 20 to 30 years, though some individuals may have continued to grow slowly for several additional years.17
Klein and Sander's 2008 study of ontogenetic stages in sauropod bone histology identified a consistent pattern across multiple sauropod taxa, including Diplodocus: rapid fibrolamellar growth during the juvenile and subadult stages, followed by a transition to more slowly deposited lamellar-zonal bone in adults as growth decelerated. The spacing of LAGs suggests that Diplodocus experienced its most rapid growth during the first 10 to 15 years of life, with growth rates declining sharply thereafter. Sellers and colleagues' 2012 digital body mass reconstruction of D. carnegii, which estimated an adult body mass of approximately 15,200 kilograms using laser-scanned skeletal data, implies that the animal needed to gain approximately 500 to 1,000 kilograms per year during its peak growth phase to achieve this mass within the timeframe indicated by bone histology.13, 15
Sander and colleagues' 2011 comprehensive review of sauropod gigantism identified several key biological innovations that enabled sauropods to achieve body sizes far exceeding those of any other terrestrial vertebrates. These include a high basal metabolic rate (inferred from fibrolamellar bone), rapid growth to reproductive maturity, a bird-like respiratory system with unidirectional airflow through the lungs (enabling efficient gas exchange at large body sizes), relatively small heads that did not require massive neck muscles for support, and the absence of mastication (chewing), which eliminated the need for a large, heavy skull with elaborate jaw musculature. Diplodocus exemplifies all of these features and serves as one of the primary model organisms for understanding the evolution of sauropod gigantism.12
The relationship between body size and metabolic rate in Diplodocus and other sauropods has been a topic of considerable debate. Traditional interpretations treated sauropods as ectothermic ("cold-blooded") animals that achieved large body sizes through the thermal inertia of gigantothermy, in which the enormous body mass maintained a stable core temperature regardless of metabolic rate. However, the fibrolamellar bone histology, rapid growth rates, and bird-like respiratory system inferred for Diplodocus all point toward an elevated metabolic rate more consistent with endothermy or some intermediate metabolic strategy. The most current view, synthesised by Sander and colleagues, is that sauropods were likely mesotherms or basal endotherms — animals with metabolic rates higher than those of modern reptiles but potentially lower than those of modern mammals, achieving their enormous sizes through a unique combination of high growth rates, efficient respiration, and reproductive strategies that prioritised quantity over parental investment.12
The Morrison Formation in which Diplodocus lived records a vast, semi-arid landscape spanning much of the western interior of North America during the Late Jurassic, approximately 155 to 148 million years ago. The formation was deposited in a mosaic of river floodplains, seasonal lakes, and open woodland, with vegetation dominated by conifers, ferns, cycads, and ginkgoes. The climate was seasonal, with pronounced dry periods evidenced by calcretes and evaporite minerals in the sediments, and the flora lacked the angiosperms (flowering plants) that would come to dominate terrestrial ecosystems during the Cretaceous. In this setting, Diplodocus and its fellow sauropods formed the ecological backbone of the herbivore community, processing enormous quantities of vegetation and likely playing a significant role in shaping the structure of the Jurassic forests they inhabited, much as elephants shape modern African savanna ecosystems through their feeding activities.10, 14
The extinction of Diplodocus and the broader diplodocid radiation at or near the Jurassic-Cretaceous boundary remains poorly understood. Diplodocids are among the most abundant and diverse sauropods in the Late Jurassic but disappear from the fossil record by the earliest Cretaceous, replaced as the dominant sauropods by titanosauriforms and macronarians. Whether this replacement reflects a mass extinction event at the Jurassic-Cretaceous boundary, a gradual competitive displacement, or simply a change in the depositional environments that preserved sauropod fossils is unclear. The Morrison Formation itself ceases deposition near the end of the Jurassic, and the paucity of earliest Cretaceous terrestrial deposits in North America creates a sampling gap that obscures the timing and mechanism of the diplodocid decline. What is clear is that the ecological role filled by Diplodocus and its relatives — as large-bodied, low-browsing herbivores processing enormous quantities of vegetation in semi-arid environments — was never precisely replicated by the sauropods that succeeded them, making the diplodocid radiation a unique chapter in the history of terrestrial herbivory.14, 16
References
Osteology of Diplodocus (Marsh) with special reference to the restoration of the skeleton in the Carnegie Museum
A reevaluation of Seismosaurus hallorum, a Late Jurassic sauropod dinosaur from New Mexico
A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda)