Overview
- Hallucigenia is a small, spiny lobopodian from the Cambrian period, first described from the Burgess Shale of British Columbia and now recognized as a stem-group onychophoran (velvet worm), placing it on the evolutionary lineage leading to one of the most ancient living animal groups.
- The reconstruction history of Hallucigenia is one of the most famous in paleontology: Simon Conway Morris's 1977 interpretation depicted the animal upside down (walking on its spines with tentacles waving above), and it was not until 1991 that Lars Ramsköld and Hou Xianguang demonstrated the correct orientation, with legs below and paired dorsal spines serving as defensive armor.
- In 2015, Martin Smith and Jean-Bernard Caron resolved the final mystery of Hallucigenia's anatomy by identifying a small, rounded head bearing a pair of simple eyes and a pharynx lined with needle-like teeth, a throat armature shared with modern velvet worms and tardigrades that supports a close evolutionary relationship among these groups within the superphylum Ecdysozoa.
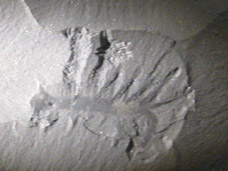
Hallucigenia is a genus of small, spiny, worm-like animals from the Cambrian period, approximately 508 million years ago, known primarily from the Burgess Shale of British Columbia, Canada, and the Chengjiang biota of Yunnan Province, China.1, 3, 10 The genus name, coined by Simon Conway Morris in 1977, reflects the "bizarre and dream-like quality" of the animal's appearance, which he found so alien that it seemed to belong to a hallucination rather than to the real history of life.1 Hallucigenia has become one of the most iconic organisms of the Cambrian explosion, famous not only for its strange form but for the extraordinary sequence of misinterpretations that plagued its reconstruction for decades, during which it was depicted upside down, backwards, or both, before its true anatomy was finally resolved.1, 3, 4 Modern analysis has established Hallucigenia as a lobopodian, a member of the broader group of "worms with legs" that are now recognized as stem-lineage panarthropods closely related to the ancestors of modern velvet worms (Onychophora).5, 8
{kind=link}
Discovery and naming
The fossils now assigned to Hallucigenia were first collected by Charles Doolittle Walcott during his extensive excavations of the Burgess Shale between 1909 and 1924, but Walcott did not recognize them as a distinct organism, tentatively grouping them with polychaete worms.1, 12 It was not until the systematic re-examination of the Burgess Shale fauna initiated by Harry Whittington in the 1960s and 1970s that the fossils received detailed study. Conway Morris, then a graduate student at Cambridge working under Whittington's supervision, published the first formal description of the genus in 1977 as Hallucigenia sparsa.1
Conway Morris described Hallucigenia as a small animal, typically 10 to 35 millimeters in body length, with an elongated, roughly cylindrical trunk bearing two rows of structures along its length: a row of seven pairs of rigid spines on one side, and a row of seven flexible, tentacle-like appendages on the other.1 One end of the body bore what appeared to be a rounded, bulbous structure, and the other end tapered to a thin tail. The organism defied placement in any known animal group, and Conway Morris was unable to determine which end was the head, which side was up, or how the animal had lived.1 In his initial reconstruction, he interpreted the rigid spines as stilt-like legs on which the animal walked, and the flexible appendages as a row of dorsal tentacles used for feeding or gas exchange.1, 2 The bulbous end was placed at the rear as a possible tail structure. This interpretation, which depicted Hallucigenia as walking on its spines with tentacles waving above its back, became one of the most reproduced images in paleontological literature and popular science.6
The great inversion
The upside-down reconstruction of Hallucigenia endured for 14 years before being overturned by new evidence from China. In 1991, Lars Ramsköld of Uppsala University and Hou Xianguang of the Yunnan Institute of Geological Sciences published a study that combined new specimens of Hallucigenia fortis from the Early Cambrian Chengjiang biota with a reanalysis of the Burgess Shale material.3 The Chinese specimens preserved the animal in a different orientation and with greater detail than the Burgess Shale fossils, and they revealed a critical feature that the original Burgess Shale specimens had obscured: the flexible appendages ended in paired claws, while the rigid spines were arranged in pairs along the dorsal surface.3
Ramsköld and Hou demonstrated that Conway Morris's reconstruction was inverted. The flexible, claw-bearing appendages were not tentacles but legs, and the rigid spines were not locomotory stilts but dorsal defensive armor.3 The corrected animal walked on its legs (seven or eight pairs, depending on the species) and bore paired rows of protective spines along its back, a body plan that made immediate functional sense: the spines would have deterred predators while the clawed legs allowed the animal to walk along the sea floor.3 The original Burgess Shale fossils had preserved the animals in a lateral view that made it ambiguous which row of structures was dorsal and which was ventral, and the paired nature of the spines had been obscured by the compression of the three-dimensional body into a flat film of carbon on the shale surface.1, 3
The inversion of Hallucigenia became a celebrated cautionary tale about the difficulties of interpreting Cambrian fossils. Conway Morris himself acknowledged the error with characteristic candor, noting in his 1998 book The Crucible of Creation that the misidentification was understandable given the quality of the original Burgess Shale specimens but was nonetheless a reminder of how profoundly prior expectations can shape the interpretation of ambiguous evidence.7
The head and pharyngeal teeth
Even after the inversion was corrected, significant uncertainties remained about Hallucigenia's anatomy, most notably the identity of the head end. The original Burgess Shale specimens showed a darkened blob at one end of the body that had been variously interpreted as a head or as a decay artifact, and a thinner structure at the opposite end.1, 15 In 2015, Martin Smith and Jean-Bernard Caron published a definitive study of Hallucigenia's head morphology using new high-resolution imaging techniques, including electron microscopy, applied to both new and existing Burgess Shale specimens.4
Smith and Caron demonstrated that the thin end of the body, not the blob, was the true head, meaning that previous reconstructions had depicted the animal not only upside down (prior to 1991) but also backwards (prior to 2015).4 The dark blob at the opposite end was reinterpreted as decay fluids that had been squeezed out of the gut during compression, not an anatomical structure at all.4 The newly identified head was small and rounded, bearing a pair of simple eyes and, most remarkably, a pharynx (throat) lined with multiple rows of fine, needle-like teeth arranged in a radial pattern around the mouth opening.4
The pharyngeal teeth of Hallucigenia proved to be of immense phylogenetic significance. Smith and Caron demonstrated that a nearly identical throat armature is found in modern velvet worms (Onychophora), which possess rows of internal jaw-like teeth derived from hardened cuticle, and in tardigrades (water bears), whose pharyngeal apparatus includes a stylet-lined buccal tube.4 The presence of pharyngeal teeth in Hallucigenia, onychophorans, and tardigrades strongly supports the hypothesis that these three groups share a common ancestor within the superphylum Ecdysozoa (molting animals), and that the toothed pharynx is an ancestral feature of the lineage that was subsequently lost in arthropods as they evolved external mouthparts.4, 5
Phylogenetic relationships
Hallucigenia is classified as a lobopodian, a grade of Cambrian animals characterized by soft, unjointed bodies with paired appendages (lobopods), which collectively represent the stem lineage of Panarthropoda, the superphylum that includes arthropods, onychophorans, and tardigrades.5, 8 Lobopodians are known from multiple Cambrian Lagerstätten including the Burgess Shale, Chengjiang, and Sirius Passet, and they display a remarkable range of body plans, from armored forms like Hallucigenia to smooth, worm-like forms such as Aysheaia.8, 11
Within the lobopodians, Hallucigenia has been placed in a clade closely related to the living Onychophora. In 2014, Smith and Javier Ortega-Hernández published a study of Hallucigenia's claw morphology, demonstrating that the claws of Hallucigenia share a distinctive layered internal structure with the claws of modern velvet worms, a shared derived character (synapomorphy) supporting their close phylogenetic relationship.5 Smith and Ortega-Hernández proposed the clade Tactopoda to include Hallucigenia, onychophorans, and tardigrades, united by this shared claw construction and by their pharyngeal armature, distinguishing them from the arthropod lineage which evolved a fundamentally different appendage structure with hardened, jointed exoskeletons.5
Ortega-Hernández's broader analysis of cephalization in panarthropods further supported Hallucigenia's position as a stem-group onychophoran, demonstrating that the simple head organization of Hallucigenia, with its paired eyes and terminal mouth, represents a plesiomorphic (ancestrally retained) condition from which the more elaborate head structures of arthropods, with their compound eyes and segmented appendages, subsequently evolved.11 The implication is that the common ancestor of all panarthropods was a small, soft-bodied, lobopodian-like animal not unlike Hallucigenia, and that the armored exoskeletons, jointed legs, and compound eyes of arthropods are derived innovations that evolved within one branch of this ancestral body plan.8, 11
Species and distribution
Two species of Hallucigenia are widely recognized. The type species, Hallucigenia sparsa, is known from the Middle Cambrian Burgess Shale (approximately 508 million years old) and is represented by more than 100 specimens in the collections of the Royal Ontario Museum and the Smithsonian Institution.1, 15 H. sparsa is typically 10 to 35 millimeters long and possesses seven pairs of legs and seven pairs of dorsal spines, with an additional pair of elongated appendages at the posterior end that may have served a sensory function.1, 15 A detailed restudy of the species by Caron and colleagues in 2013, based on over 100 new and existing specimens, confirmed the basic body plan while revealing new anatomical details including the gut trace, the relative sizes of the legs and spines, and population-level variation in body proportions.15
The second species, Hallucigenia fortis, is known from the Early Cambrian Chengjiang biota of Yunnan Province, China (approximately 518 million years old), and is both larger and slightly different in proportions from the Burgess Shale species.3, 10 H. fortis can reach up to 50 millimeters in length and possesses more robust spines relative to its body size than H. sparsa.3 It was the superior preservation of H. fortis specimens, which in some cases retained three-dimensional structure rather than being completely flattened, that enabled Ramsköld and Hou to overturn the original inverted reconstruction.3
Additional lobopodians sharing features with Hallucigenia have been described from Cambrian deposits worldwide, including Microdictyon (which bore mineralized sclerite plates rather than spines) and Onychodictyon from Chengjiang.8, 10 These related forms demonstrate that armored lobopodians were a diverse and widespread component of Cambrian marine faunas, not an isolated oddity confined to a single locality.8
Significance for understanding early animal evolution
Hallucigenia occupies a unique place in the history of paleontology and in the scientific understanding of early animal evolution. Its reconstruction history, from upside down to right side up to finally correct head-to-tail orientation, has become a standard teaching example of how science self-corrects through the accumulation of new evidence and the willingness to discard established interpretations when they are contradicted by data.1, 3, 4, 7 More substantively, the phylogenetic placement of Hallucigenia as a stem-group onychophoran has profoundly shaped understanding of the Cambrian explosion by demonstrating that at least some of the apparently "bizarre" organisms of the Burgess Shale and Chengjiang are not members of extinct phyla with no living descendants, as Gould argued in Wonderful Life, but rather stem-group members of extant animal lineages.5, 6, 7
The pharyngeal teeth of Hallucigenia provide a concrete example of how Cambrian fossils can illuminate the deep evolutionary relationships among living animal groups. The discovery that Hallucigenia, onychophorans, and tardigrades share a toothed pharynx would have been impossible to predict from the study of living organisms alone, because arthropods, the most diverse and ecologically dominant panarthropods, have lost this feature entirely.4 Only by examining the fossil record of the Cambrian, when the stem lineages of these groups were still recognizable as distinct from their crown groups, could this ancestral character be identified and the evolutionary history of panarthropod mouthparts reconstructed.4, 14
In this way, Hallucigenia exemplifies the irreplaceable role of Cambrian Lagerstätten in reconstructing the tree of life. Without the exceptional preservation of soft-bodied organisms in the Burgess Shale and Chengjiang, the evolutionary connections between living velvet worms, tardigrades, and arthropods would remain far more obscure, and the deep history of these groups, which together account for the vast majority of animal species on Earth, would be largely unknowable.4, 5, 8
References
New organisms from the Middle Cambrian of the Burgess Shale and the role of the Middle Cambrian in the Cambrian explosion
Cambrian lobopodians and extant onychophorans provide new insights into early cephalization in Panarthropoda
The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group
A large new leanchoiliid from the Burgess Shale and the influence of inapplicable characters on stem arthropod phylogeny