Overview
- The family Equidae preserves one of the most complete evolutionary sequences in the entire fossil record, tracing 55 million years of continuous anatomical change from Hyracotherium, a small forest-dwelling browser with four toes on the front feet, through a succession of increasingly large, long-legged, high-toothed forms, to the single-hoofed modern horse Equus.
- The trajectory is not a straight ladder but a richly branching bush: dozens of equid lineages coexisted and went extinct at various points, and body size reversed direction multiple times, but the overall trends toward toe reduction, enlarged hypsodontic molars, and expanded brain volume are robustly documented across hundreds of specimens from North American, European, and Asian deposits.
- Horses originated in North America, dispersed across Beringia and the Central American land bridge to colonize every continent except Antarctica and Australia, then went extinct on their continent of origin around 10,000 years ago — an extinction reversed only when Spanish conquistadors reintroduced domesticated Equus caballus in the sixteenth century.
No lineage in the entire fossil record has been studied more intensively or cited more frequently as evidence for evolution than the family Equidae. From the moment Othniel Charles Marsh displayed the North American horse sequence before Thomas Henry Huxley in 1876, Equidae became the iconic demonstration that species are not fixed entities but mutable populations changing continuously through geological time. The sequence runs from Hyracotherium in the early Eocene, approximately 55 million years ago, to Equus in the Pleistocene and present — a span of more than 50 million years, represented by hundreds of specimens from dozens of localities across North America, Europe, and Asia, preserving skeletal transformations so finely graded that the evolutionary transitions are visible in single museum drawers.1, 2
._The_origin_and_evolution_of_life,_on_the_theory_of_action,_reaction_and_interaction_of_energy.jpg){kind=link}
The history of the group has also served as a corrective to naive progressivism in evolutionary thinking. Mid-twentieth-century textbooks presented horse evolution as a single straight line of ancestors and descendants, a ladder of increasing size and sophistication marching inevitably toward the modern thoroughbred. Subsequent work overturned this reading entirely. Equidae is a richly branching phylogenetic bush in which dozens of lineages coexisted at any one time, body size both increased and decreased multiple times independently, and the vast majority of equid diversity ended in extinction with no living descendants. The story that emerges from a century and a half of collecting is more interesting, and more genuinely evolutionary, than the ladder ever was.1, 2, 11
Hyracotherium and the Eocene origin
Hyracotherium leporinum was first described by Richard Owen in 1840 from fragmentary British material, and for decades the animal was known by the popular name “Eohippus” (dawn horse) coined by Othniel Charles Marsh for closely related North American specimens. Modern taxonomy treats “Eohippus” as synonymous with Hyracotherium or as a closely allied genus, and the International Code of Zoological Nomenclature gives priority to Owen’s earlier name. Phylogenetically, Hyracotherium sits at or near the base of Perissodactyla, the order that also includes tapirs and rhinoceroses, and represents the ancestral condition from which all equids are derived.1, 8
Hyracotherium was a small animal, roughly the size of a fox terrier, standing approximately 20 to 45 centimetres at the shoulder and weighing an estimated 5 to 20 kilograms depending on species. Its limbs were slender but retained a relatively generalized mammalian proportions: the forefeet bore four functional toes (with vestigial remnants of a fifth), and the hindfeet bore three. Each digit terminated not in a true hoof but in a small, hoof-like pad, and the foot posture was digitigrade — the animal walked on its toes rather than the full sole of the foot, but the weight was distributed across several digits rather than concentrated on one. The teeth were brachydont (low-crowned), with rounded, bunodont cusps well suited for crushing soft leaves, fruits, and other vegetation in the closed-canopy forest environments that dominated the early Eocene across North America and Europe, when the two continents were separated by only a shallow seaway and the climate was substantially warmer and wetter than today.1, 2, 14
The brain of Hyracotherium was proportionally smaller than that of modern Equus, with relatively modest temporal and frontal lobes, consistent with the generally small encephalization quotients of early Eocene mammals. The orbit was positioned more anteriorly on the skull, the facial region was longer relative to the cranial vault, and the overall skull shape resembles the generalized perissodactyl ancestor more than it does any modern horse.1
Oligocene and Miocene diversification
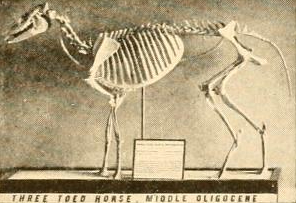
Through the Eocene and into the Oligocene, equid lineages diversified substantially in North America while simultaneously dispersing to Europe and Asia via the land connections that episodically connected the continents across Beringia. Mesohippus, which flourished during the Oligocene approximately 37 to 32 million years ago, represents a significant advance over Hyracotherium in almost every anatomical dimension. It stood approximately 60 centimetres at the shoulder, had three functional toes on both the front and hind feet (though a vestigial splint bone on the forefoot records the former fourth digit), and possessed slightly higher-crowned teeth with more complex enamel folding on the molars. The facial region was elongated, expanding the space available for larger molars and creating the longer muzzle characteristic of later horses.1, 2
Parahippus, from the early Miocene approximately 23 to 17 million years ago, represents the transition between the browsing horses and the grazing horses that would come to dominate the later Cenozoic. Its molars show incipient hypsodonty — the teeth were slightly taller-crowned than those of earlier equids, and the enamel cementoenamel junction was beginning to move higher up the tooth, a geometry that exposes more enamel surface area for grinding abrasive foodstuffs while also extending the functional lifespan of the tooth by providing a larger reserve of crown material above the gumline. Microwear analysis of Parahippus teeth reveals a mixed diet of browse and grass, consistent with its temporal position as the savanna ecosystems that would define North American midcontinental environments for the rest of the Cenozoic began to expand during the global cooling and drying trend of the middle Miocene.4, 12, 13
Merychippus, the dominant equid of the middle to late Miocene approximately 17 to 11 million years ago, marks the decisive shift to obligate grazing. Its molars are fully hypsodont — the crowns are tall, the enamel is folded into complex patterns of ridges and lakes visible on the occlusal surface, and cement fills the infundibula between the enamel folds, creating a grinding surface of heterogeneous hardness that wears to maintain a rough, file-like texture throughout the tooth’s life. This dental architecture is a direct response to the mechanical and chemical properties of grass: unlike most browse, grass incorporates silica phytoliths into its cell walls, which abrade teeth far more rapidly than soft dicotyledonous foliage, and the longer a tooth crown remains above the gumline, the longer it can sustain the demands of a grass diet.4, 12, 13
Merychippus retained three toes on each foot, but the lateral digits had been substantially reduced: the central third toe was considerably larger and more robust than the second and fourth, bore a true hoof, and carried nearly all of the animal’s weight. The side toes barely touched the ground and would have had minimal locomotor function on firm substrates, functioning primarily as splints and perhaps providing some stability on soft ground. Body size had increased to approximately 90 to 100 centimetres at the shoulder. Several distinct lineages descended from Merychippus or its close relatives, radiating into a diversity of ecological niches across the expanding grasslands of North America during the late Miocene — a period when equid diversity reached its peak, with as many as a dozen genera coexisting on the same continent.1, 2, 3
Key equid genera through geological time1, 2
| Genus | Epoch | Age (Ma) | Toes (front / rear) | Tooth type | Est. shoulder height |
|---|---|---|---|---|---|
| Hyracotherium | Early Eocene | 55–45 | 4 / 3 | Brachydont, bunodont | 20–45 cm |
| Mesohippus | Oligocene | 37–32 | 3 / 3 | Low hypsodont | ~60 cm |
| Parahippus | Early Miocene | 23–17 | 3 / 3 | Transitional hypsodont | ~75 cm |
| Merychippus | Middle–Late Miocene | 17–11 | 3 / 3 (central dominant) | Fully hypsodont | ~100 cm |
| Pliohippus | Pliocene | 12–6 | 1 / 1 (vestigial splints) | High hypsodont | ~120 cm |
| Equus | Pleistocene–present | 4–0 | 1 / 1 | Very high hypsodont | 140–165 cm |
Pliohippus, Equus, and the completion of toe reduction
Pliohippus, ranging from approximately 12 to 6 million years ago through the Miocene and into the early Pliocene, is the first equid genus to have achieved effectively single-toed locomotion. The lateral digits were reduced to vestigial splint bones embedded in connective tissue along the cannon bone (the enlarged and fused central metapodial), leaving no external trace on the living animal. The single hoof bore the entire weight of the body, the limbs were proportionally longer and more slender than those of any previous equid, and the overall body form was approaching that of a small modern horse.1, 3 Long treated as the direct ancestor of Equus, Pliohippus has been re-evaluated by cladistic analyses as a side branch; the genus most directly ancestral to Equus is now generally considered to be Dinohippus, a contemporary of Pliohippus from the late Miocene of North America that shares more derived features of the skull and teeth with the modern genus.2, 3
The genus Equus appears in the North American fossil record approximately 4 million years ago and diversified rapidly through the Pliocene and Pleistocene into numerous species distributed across multiple continents. Living representatives of the genus include the domestic horse (Equus caballus), the Przewalski’s horse (E. przewalskii), three species of zebra, the African wild ass (E. africanus), the onager (E. hemionus), and the kiang (E. kiang). Extinct Pleistocene Equus species populated North America, Europe, Asia, Africa, and South America in impressive diversity, with some North American forms reaching body sizes comparable to the largest modern draft breeds.1, 7, 10
The anatomical differences between Equus and the earliest equids are substantial: body mass has increased by perhaps a factor of 60 to 100, tooth crown height has increased by a similar magnitude relative to crown width, the number of functional digits has decreased from four (front) and three (rear) to one on each foot, and the brain volume relative to body mass has roughly doubled. Yet these changes accumulated gradually, not saltatorially, across a succession of intermediate forms that document each stage of the transformation.1, 2
A branching bush, not a ladder
The image of horse evolution as a straight progressive sequence owes its persistence partly to the way museum displays were historically arranged and partly to the genuine directionality of certain trends. In reality, equid phylogeny is characterized by a degree of diversity and complexity that the ladder metaphor completely obscures. During the Miocene, three to five equid tribes coexisted in North America simultaneously, representing different combinations of dental and locomotor adaptations, different body sizes, and different dietary specializations. Archaeohippus, a Miocene equid contemporary with Merychippus, actually reduced its body size relative to its Oligocene ancestors, a clear reversal of the supposed general trend. Nannippus, a lineage of small three-toed horses that persisted into the Pliocene alongside the large single-toed forms, demonstrates that reduction in toe number and increase in body size were not obligately linked and could evolve at different rates in different lineages.1, 2, 11
The correct picture, firmly established by phylogenetic analyses using both morphological and molecular data, is a tree with a single stem rooted in early Eocene North America, branching repeatedly through the Eocene, Oligocene, Miocene, and Pliocene, with the vast majority of branches ending in extinction and only the single lineage leading to Equus surviving to the present. Body size evolution within this tree, analyzed in a phylogenetic framework, shows multiple independent increases and at least several decreases, consistent with a pattern of Cope’s rule operating irregularly across the clade rather than as an iron constraint.2, 11 The directional trends that are real — hypsodonty increasing in most grazing lineages, digit number decreasing in the main line leading to Equus — reflect consistent selective pressures imposed by expanding grassland environments, not some intrinsic drive toward a predetermined end point.
Understanding equid evolution as a bush also explains a fact that puzzled earlier workers: the fossil record contains many equids that do not fit neatly into any simple ancestor-descendant sequence. These animals are not anomalies to be explained away. They are the expected result of a clade diversifying under ecological and climatic pressures that drove parallel evolution of similar adaptations in multiple independent lineages. The acquisition of hypsodonty, for example, occurred in at least three distinct equid lineages during the Miocene, each responding independently to the same dietary challenge posed by abrasive grasses.4, 13
Molecular phylogenetics and confirmation
The anatomical sequence established by paleontologists has been independently confirmed and extended by molecular data. Perissodactyla, the order containing horses, tapirs, and rhinoceroses, is placed by genomic analyses as the sister group of Cetartiodactyla (whales, pigs, cattle, deer, and their relatives), together forming a clade sometimes called Laurasiatheria along with bats, carnivorans, and pangolins. Within Perissodactyla, Equidae and Tapiridae are consistently resolved as more closely related to each other than either is to Rhinocerotidae, though all three families share a common perissodactyl ancestor in the early Eocene.8, 9
The complete sequencing of the horse genome, published in 2009, confirmed the fundamental mammalian relationships and also provided a detailed internal framework for equid phylogeny. Comparisons among living Equus species place the divergence of the horse and donkey lineages at approximately 4 to 4.5 million years ago, consistent with the first appearance of Equus in the fossil record.9 Ancient DNA extracted from late Pleistocene equid fossils has allowed molecular phylogenetics to reach directly into the fossil record, confirming the stratigraphic sequence while adding resolution to the internal branching order of late Pleistocene Equus populations. Analyses of permafrost-preserved horse specimens from Alaska, the Yukon, and Siberia have demonstrated that the North American Pleistocene horse populations were distinct from the Eurasian populations that gave rise to the domesticated horse and Przewalski’s horse, and that the New World horses went extinct as a discrete clade rather than as a subset of a continuously distributed global population.5, 6, 10
The congruence between the fossil sequence and the molecular phylogeny is striking. The two lines of evidence were developed largely independently — the paleontological sequence using morphological comparisons of hundreds of fossil specimens, the molecular phylogeny using sequence data from living and ancient DNA — yet they converge on the same branching topology, the same relative timing of divergence events, and the same geographic origin for the group. This consilience of independent methods reaching the same conclusion is among the most powerful forms of evidence available in science.8, 9
North American origin and global dispersal
Equidae originated in North America, and for the first 45 million years of the family’s existence, the continent served as the primary theater of equid evolution. The rich sequence of North American equid faunas documented in the White River Badlands of South Dakota, the Fayum equivalent formations of Nebraska’s Ogallala Group, and the Pleistocene tar pits of California reflects tens of millions of years of in situ diversification on a single landmass. Equids periodically dispersed to other continents when Beringia, the land connection between Alaska and Siberia, was exposed during periods of low sea level associated with glacial maxima: ancestors of the Old World horses crossed into Eurasia during the Miocene, where they gave rise to additional lineages, and Equus itself dispersed from North America to Eurasia, Africa, and South America during the Pleistocene.1, 2, 14
The dispersal of Equus to South America occurred approximately 1.5 to 1.0 million years ago, following the formation of the Isthmus of Panama approximately 3 million years ago. South American Equus species are among the many large mammals that arrived from North America during the Great American Biotic Interchange and subsequently diversified on the southern continent, only to go extinct at the end of the Pleistocene along with the megafaunal communities of which they were part.1, 10
The extinction of horses in North America at approximately 10,000 years ago is one of the defining events of the late Pleistocene megafaunal extinctions. The cause or causes remain actively debated: candidates include the hunting pressure exerted by the Clovis and pre-Clovis human populations that colonized the Americas during the terminal Pleistocene, the rapid climate shifts accompanying the end of the last glacial maximum, or a combination of both. Ancient DNA evidence indicates that North American horse populations experienced significant genetic diversity loss in the millennia before final extinction, consistent with a shrinking population under sustained pressure rather than a sudden catastrophe.7, 10 Whatever the cause, the result was the complete elimination of the family that had originated and diversified on that continent over 50 million years, leaving no horses anywhere in the Americas for approximately 9,000 years.
The reintroduction of horses to the Americas began with the second voyage of Christopher Columbus in 1493 and accelerated with the Spanish conquest of the mainland in the early sixteenth century. Equus caballus recolonized its ancestral continent with remarkable speed: escaped and released domestic horses established feral populations, of which the North American mustang is the most celebrated, within decades of the initial introduction. The horses encountered by Plains Indian peoples in the eighteenth century were not a new phenomenon from their perspective but rather the return of an animal whose fossil ancestors lay in the ground beneath their feet, an irony whose geological dimension was not fully appreciated until the nineteenth-century fossil discoveries made the connection explicit.6, 10
The sequence as evidence for evolution
The Equidae fossil record has occupied a central place in discussions of evolutionary evidence since Marsh presented his sequence to Huxley, and Huxley immediately used it as the centerpiece of his New York lecture on evolution in 1876. The sequence is compelling for reasons that go beyond the sheer number of specimens. The anatomical changes documented are not trivial variations on a single body plan but fundamental reorganizations of the locomotor apparatus, the dentition, and the skull. The reduction from four functional toes to one involves a series of changes in the relative proportions and robust of the metatarsals and phalanges that are mechanically coherent: at each stage, the body weight is supported by a skeletal arrangement that functions, and each successive reduction in the lateral digits involves compensatory strengthening of the central digit and elongation of the metapodial that maintains locomotor efficiency.1, 15
The geographic distribution of the sequence provides additional evidentiary weight. The oldest equid fossils are found in North American and European early Eocene deposits. Progressively more derived equids appear in progressively younger stratigraphic levels across the continent, with the spatial and temporal pattern of first appearances consistent with in situ evolution and episodic dispersal rather than independent origin in multiple locations. No anatomically modern Equus has ever been found in Eocene rocks, and no Hyracotherium-grade equid has ever been found in Pleistocene deposits. The temporal ordering of the sequence is not a post-hoc arrangement imposed by the researcher but is determined independently by the radiometric ages of the containing sediments, stratigraphy, and magnetostratigraphy, all of which confirm the same relative sequence.1, 2
The horse sequence also illustrates the explanatory power of understanding evolution as a response to changing environmental conditions rather than as an internal program of improvement. The shift from brachydont to hypsodont dentition tracks, with remarkable fidelity in the fossil record, the expansion of grassland ecosystems during the middle Miocene global cooling and drying. This correlation was first noted qualitatively and has since been confirmed quantitatively: tooth crown height indices across North American equid faunas increase in step with independently derived estimates of grass cover based on phytolith assemblages in sediments of the same age. The teeth did not become taller because horses were “progressing” but because taller teeth conferred a survival advantage in an environment where soft browse was progressively replaced by abrasive grass.4, 12, 13
Creationist arguments that the horse sequence is fabricated or misrepresented have been comprehensively addressed in the scientific literature. The claim that the sequence consists of unrelated animals arranged arbitrarily is refuted by the cladistic analyses that recover the same branching order from multiple independent anatomical character systems, by the stratigraphic ordering of the fossils, and by the molecular phylogenies derived from living and ancient DNA. The claim that body size increases monotonically, and is therefore too regular to be the product of natural selection, is refuted by the documented cases of body size reversal within equid lineages and by the well-understood population genetic mechanisms by which directional selection can produce sustained trends over millions of generations. The sequence is, in the judgment of systematic paleontology, one of the most thoroughly verified evolutionary sequences known to science, and the convergence of multiple independent methods on the same phylogenetic and temporal reconstruction leaves no scientifically credible alternative to common descent as its explanation.1, 2, 8
References
- MacFadden, B. J. · Fossil horses: systematics, paleobiology, and evolution of the family Equidae. Cambridge University Press, 1992.
- MacFadden, B. J. · Horses and their relatives: a review of the evolution of the family Equidae. Annual Review of Earth and Planetary Sciences 33: 147–175, 2005.
- Prado, J. L. & Alberdi, M. T. · Explosive radiation of Mio-Pliocene horses in the New World with the origin of the one-toed grazing horses. Journal of Vertebrate Paleontology 24: 660–676, 2004.
- Jardine, P. E., Janis, C. M., Sahney, S. & Benton, M. J. · Hypsodonty in ungulates: an adaptation for grass consumption or for rapid growth rates? Palaeogeography, Palaeoclimatology, Palaeoecology 321–322: 98–110, 2012.
- Palkopoulou, E. et al. · A comprehensive genomic history of extinct and living elephants. Proceedings of the National Academy of Sciences 115: E2566–E2574, 2018.
- Schubert, M. et al. · Genomic analysis of the extinct Eurasian wild horse and the origin of domestication. Nature Genetics 46: 1330–1336, 2014.
- Librado, P. et al. · Ancient genomics and the origin of horse domestication in the Eurasian steppes. Nature 598: 634–640, 2021.
- Guthrie, R. D. · Rapid body size decline in Alaskan Pleistocene horses before extinction. Nature 426: 169–171, 2003.
- Steiner, C. C. & Ryder, O. A. · The phylogenetic position of the horse lineage: evidence from complete mitochondrial genome sequences of extant perissodactyls. Zoological Journal of the Linnean Society 163: 1289–1303, 2011.
- Wade, C. M. et al. · Whole-genome sequencing of the horse (Equus caballus). Science 326: 865–867, 2009.
- Heintzman, P. D. et al. · Spread of horses in the Americas: new genomic and archaeological evidence. Current Biology 28: 2892–2897, 2018.
- Alroy, J. · Cope’s rule and the dynamics of body mass evolution in North American fossil mammals. Science 280: 731–734, 1998.
- Strömberg, C. A. E. · Grass phytoliths as a dietary indicator in large mammalian herbivores: evidence from modern and fossil teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 207: 209–246, 2004.
- Mihlbachler, M. C., Rivals, F., Solounias, N. & Semprebon, G. M. · Evolution of hypsodonty in equids: testing a hypothesis of climate-driven change. Science 331: 1178–1181, 2011.
- Janis, C. M. · The antiquity and biogeography of Cenozoic mammals in North America. In: Evolution of Tertiary Mammals of North America, Cambridge University Press, 1998.
- Coombs, M. C. · Locomotion in fossil horses and the origin of perissodactyls. Paleobiology 4: 246–258, 1978.
References
Explosive radiation of Mio-Pliocene horses in the New World with the origin of the one-toed grazing horses
Hypsodonty in ungulates: an adaptation for grass consumption or for rapid growth rates?
The phylogenetic position of the horse lineage: evidence from complete mitochondrial genome sequences of extant perissodactyls
Grass phytoliths as a dietary indicator in large mammalian herbivores: evidence from modern and fossil teeth