Overview
- Paraceratherium was a genus of giant hornless rhinocerotoid that inhabited Eurasia during the Oligocene epoch from roughly 34 to 23 million years ago, and is widely regarded as the largest land mammal in the fossil record, with the best-preserved specimens reaching shoulder heights near 4.8 metres and body masses around 17 tonnes.
- The taxonomic history of the genus is unusually tangled because giant rhinocerotoid remains discovered between the late nineteenth century and the 1930s in Pakistan, Mongolia, Kazakhstan and China were named under at least eight separate genera — including Baluchitherium, Indricotherium, Aralotherium and Dzungariotherium — before the 1989 revision by Lucas and Sobus consolidated almost all valid species under the senior name Paraceratherium.
- New skeletal material from the Linxia Basin of China described in 2021 added Paraceratherium linxiaense to the genus and demonstrated that giant rhinos dispersed across the Tibetan region during the late Oligocene, indicating that the future Tibetan Plateau had not yet risen as a barrier to large-mammal movement and reshaping prior reconstructions of Asian Cenozoic biogeography.
Paraceratherium is an extinct genus of giant hornless rhinocerotoid perissodactyls that inhabited Eurasia during the Oligocene epoch, from roughly 34 to 23 million years ago. It is the largest land mammal documented in the fossil record by reasonably complete material: the best-preserved individuals approached shoulder heights of 4.8 metres, body lengths near 7.4 metres, and body masses on the order of 15 to 20 tonnes — figures that exceed every fully extant terrestrial mammal and approach the largest plausible reconstructions of Pleistocene proboscideans.6, 7 Despite the popular image of dinosaurs as the only true giants of deep time, Paraceratherium demonstrates that the post-K–Pg mammalian adaptive radiation produced terrestrial herbivores rivalling the smaller sauropods in raw body mass.10, 15

{kind=link}
The genus has had an unusually tangled history. The earliest fragmentary remains were collected from the Bugti Hills of Balochistan, Pakistan in the late nineteenth century, and the first formal taxonomic name — Aceratherium bugtiense — was applied by the British geologist Guy Ellcock Pilgrim in 1910; over the following three decades, similar giants discovered across Mongolia, Kazakhstan and China were named under separate genera including Baluchitherium, Indricotherium, Aralotherium and Dzungariotherium, only to be progressively synonymised through the twentieth century.1, 3, 5 A 1989 revision by Spencer Lucas and Jay Sobus established Paraceratherium, the oldest available name, as the senior synonym for almost all valid species, and the genus has since been refined by new Chinese material that places it within a broader Eurasian biogeographic context.5, 8
Discovery and original naming
The fossil remains now assigned to Paraceratherium were first encountered in the late nineteenth century in the Bugti Hills of what is today Balochistan, Pakistan, where the British Indian Geological Survey collected isolated teeth and limb fragments of large mammals from the Chitarwata Formation. These early finds were too fragmentary to support a confident identification, and it was not until the field seasons of 1907 and 1908 that the British geologist Guy Ellcock Pilgrim recovered the partial upper jaw, lower dentition and posterior mandible that would form the basis of the first formal description.6, 11 Pilgrim, finding no obvious distinguishing features that separated his material from the European hornless rhinoceros genus Aceratherium, named the species Aceratherium bugtiense in his 1910 paper Notices of new mammalian genera and species from the Tertiaries of India and assigned it to the late Oligocene of the Bugti beds.11, 20
In 1910, the British palaeontologist Clive Forster-Cooper led a fresh expedition to Dera Bugti and recovered substantially more complete remains of the same animal, including additional jaw material that allowed him to recognise that the lower incisors were strongly down-turned and otherwise quite unlike those of true Aceratherium. In a preliminary 1911 paper in the Annals and Magazine of Natural History, Forster-Cooper formally erected the new genus Paraceratherium — literally "near the hornless beast" — to accommodate Pilgrim's species.1 A more detailed monographic treatment followed in his 1924 paper in Philosophical Transactions of the Royal Society B, in which he described the skull and dentition of additional specimens and clarified the broad outlines of the animal's anatomy.2 Forster-Cooper subsequently coined a second name, Baluchitherium osborni, for very large limb material that he believed represented a separate, even larger giant rhinoceroid; this duplication seeded much of the taxonomic confusion that would dog the group for the next eighty years.6, 16
The early twentieth century was an unusually competitive era in vertebrate palaeontology, and the giant rhinos quickly became prizes in an international race for spectacular Asian fossils. Russian, German, French, British and American institutions all dispatched expeditions to Central Asia in the 1910s and 1920s, and each tended to coin a new generic name for whatever giant remains they recovered, with little reference to the others' material. The historian Chris Manias has documented how the resulting profusion of names — Baluchitherium, Indricotherium, Thaumastotherium, Aralotherium, Dzungariotherium and others — reflected the imperial and institutional rivalries of the period at least as much as it did genuine biological differences between the specimens.16
Taxonomic history and synonymy
The Soviet palaeontologist Aleksei Borissiak coined the name Indricotherium in 1916 for giant rhinocerotoid material from the Aral region of Kazakhstan, and Maria Pavlova added Indricotherium transouralicum in 1922 for further material from the Turgai region. The name Indricotherium derives from the Indrik, a vast horned bull of Russian folklore, and it would persist in popular and museum usage for most of the twentieth century. Roy Chapman Andrews's Central Asiatic Expeditions of 1922–1925, sponsored by the American Museum of Natural History, recovered substantial giant rhinocerotoid material from the Hsanda Gol Formation of central Mongolia, including the AMNH 18650 skull collected near Loh in 1922; Henry Fairfield Osborn named this material Baluchitherium grangeri in 1923, in honour of the field leader Walter Granger.3, 16
By the mid-1930s, several palaeontologists had begun to suspect that the proliferation of generic names did not reflect underlying anatomical reality. In 1935, Granger and Gregory published a revised skeletal restoration of the Mongolian giant in the American Museum Novitates, and in 1936 they followed up with a long Bulletin paper that re-examined nearly all the available giant rhinocerotoid material then known and concluded that the differences between Baluchitherium, Indricotherium and Paraceratherium were almost entirely artefacts of preservation and intraspecific variation.3, 4 They retained Baluchitherium as a genus largely on grounds of priority and prior usage, but their analysis essentially anticipated the modern synonymy.3
The definitive revision came more than half a century later, when Spencer Lucas and Jay Sobus published a systematic re-examination of all named indricothere taxa in 1989. Their analysis demonstrated that the supposed generic distinctions among Paraceratherium, Baluchitherium, Indricotherium, Aralotherium, Pristinotherium and several others were not supported by consistent characters and that the entire group represented a single genus. Because Paraceratherium was the senior name — published in 1911, before all the major rivals — it took priority under the International Code of Zoological Nomenclature and absorbed the others as junior synonyms.5 Lucas and Sobus recognised four valid species in the genus: P. bugtiense from the late Oligocene of Pakistan, P. transouralicum from Kazakhstan and Mongolia, P. prohorovi from Kazakhstan, and P. orgosensis from northwestern China. Subsequent work by Qiu and Wang on Chinese material refined this scheme, and the description of P. huangheense in 2017 by Li and colleagues from Gansu Province, followed by P. linxiaense in 2021, brought the currently accepted species count to between four and six, depending on whether certain disputed taxa are retained.8, 13, 18
Currently recognised species of Paraceratherium5, 8, 13, 18
| Species | Author and year | Type locality | Approximate age |
|---|---|---|---|
| P. bugtiense | (Pilgrim, 1908) | Bugti Hills, Pakistan | Late Oligocene |
| P. transouralicum | (Pavlova, 1922) | Turgai region, Kazakhstan | Late Oligocene |
| P. prohorovi | (Borissiak, 1939) | Aral region, Kazakhstan | Early Oligocene |
| P. orgosensis | (Chiu, 1973) | Junggar Basin, China | Late Oligocene |
| P. huangheense | Li et al., 2017 | Wushan Basin, Gansu, China | Late Oligocene |
| P. linxiaense | Deng et al., 2021 | Linxia Basin, Gansu, China | Late Oligocene (~26.5 Ma) |
Phylogeny and relationships
Paraceratherium belongs to the superfamily Rhinocerotoidea, the broader rhinoceros lineage within the order Perissodactyla. Its precise familial placement has shifted considerably as understanding of early rhinocerotoid relationships has improved. For most of the twentieth century the giant rhinos were treated as a distinct family, Indricotheriidae, and were sometimes considered a subfamily of Rhinocerotidae. In 1966 the American palaeontologist Leonard Radinsky reorganised early rhinocerotoids and placed the indricotheres as a subfamily, Indricotheriinae, within the Hyracodontidae — the so-called "running rhinos" of the Eocene and Oligocene — and this arrangement was followed for several decades.6
More recent phylogenetic analyses have substantially revised this picture. In 2016 Wang and colleagues described Pappaceras meiomenus, a new forstercooperiine rhinocerotoid from the early Eocene Arshanto Formation of Inner Mongolia, and used the new material to perform the first comprehensive cladistic analysis of early rhinocerotoid relationships. Their results recovered the giant rhinos as a monophyletic group within a distinct family, Paraceratheriidae, that fell outside the traditional Hyracodontidae and was actually closer to the true rhinos (Rhinocerotidae) than to the "running rhinos."9 Within Paraceratheriidae, two subfamilies are now recognised: the basal Forstercooperiinae, including small-bodied Eocene forms such as Forstercooperia and Pappaceras, and the derived Paraceratheriinae, which includes the increasingly large genera Juxia, Urtinotherium and finally Paraceratherium itself.9 The group thus represents a clear ecological and morphological trajectory toward gigantism, beginning with dog-sized Eocene browsers and culminating in the late Oligocene with the largest land mammals ever to exist.6, 9
The phylogenetic analysis published by Deng and colleagues in 2021, which incorporated the new P. linxiaense material, recovered the same broad pattern: Paraceratheriidae as a distinct rhinocerotoid family sister to Rhinocerotidae plus Amynodontidae, with the genus Paraceratherium as a relatively derived terminal branch within Paraceratheriinae.8 Within the genus, their analysis placed P. linxiaense and P. lepidum as sister taxa with a close relationship to P. bugtiense, suggesting that the Pakistani species represents a southward dispersal from a Central Asian centre of origin.8
Anatomy and skeleton
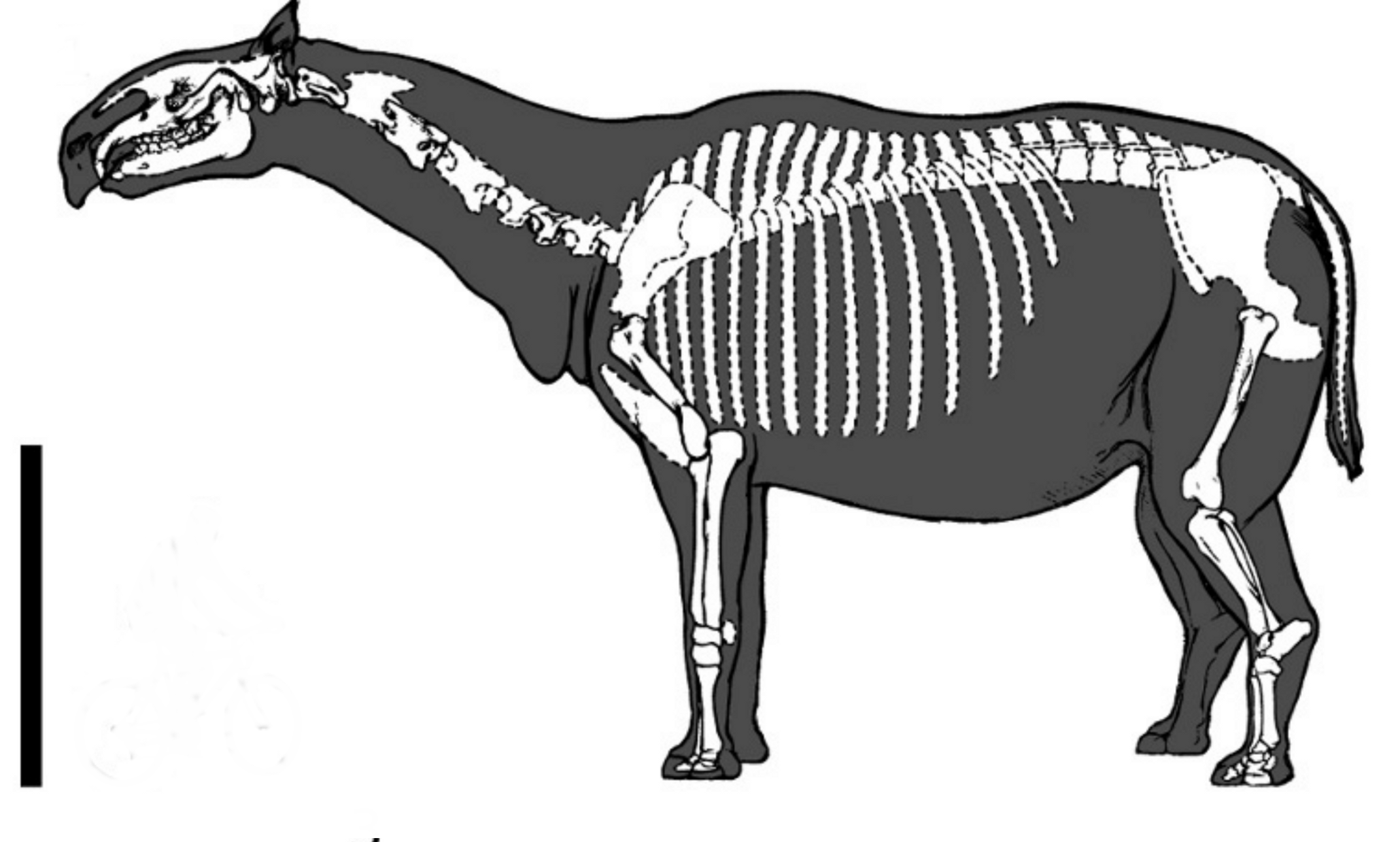
The skeleton of Paraceratherium conveys an unmistakable impression of a giraffe-bodied rhinoceros: a long, mobile neck capped by a long but relatively narrow skull, supported by a massive trunk and column-like, graviportal limbs adapted for sustained weight-bearing rather than rapid locomotion. The largest known skulls reach approximately 1.3 metres in length and bear a single pair of large, conical incisors in each jaw. The upper incisors point downward and the lower ones project forward, forming an interlocking arrangement quite unlike that of any modern rhinoceros and superficially resembling small tusks; canines are entirely absent.2, 6 The cheek teeth are low-crowned and bear a pi-shaped enamel pattern on the upper molars and an L-shaped pattern on the lowers, both characteristic of rhinocerotoids more generally and consistent with a soft-leaf browsing diet rather than the abrasive grazing diet of later horses and bovids.6
The neck of Paraceratherium is reconstructed at 2.0 to 2.5 metres in length, although a fully articulated cervical column has yet to be recovered. The atlas and axis vertebrae are conspicuously broad, and the more posterior cervicals bear large zygapophyses and elongate neural spines that together formed a pronounced shoulder hump for the attachment of the powerful nuchal ligaments and epaxial muscles needed to support the massive skull at the end of a long lever arm.2, 6 The forelimbs are columnar, with extremely robust humeri, radii and metacarpals; weight is borne primarily on the third digit, in keeping with the perissodactyl bodyplan, but the medial and lateral digits remain functional in load distribution. Limb proportions and joint mechanics indicate that Paraceratherium was effectively a slow, graviportal browser that walked rather than ran — a mode of locomotion convergent with that of large proboscideans rather than with extant rhinos, which retain considerable cursorial ability.10
{kind=link}
The shape of the nasal bones and the position of the external nares have been a recurring focus of debate. Forster-Cooper noted in 1924 that the nasal opening was retracted relative to that of typical rhinoceroses and inferred a small, prehensile snout or short proboscis comparable to that of a tapir or a black rhinoceros.2 Mark Witton has more recently argued, on comparative-anatomical grounds, that the available evidence does not unambiguously support a true trunk and that a deeply prehensile upper lip is more parsimonious; the question remains unresolved in the absence of soft-tissue impressions.6
Body size estimates
Quantifying the size of Paraceratherium is complicated by the fact that no single individual is known from a complete, articulated skeleton. Almost every published estimate has been built up by composite reconstruction, scaling isolated limb elements and vertebrae against modern proxies, and the resulting figures have varied widely. The earliest estimates, dating to the 1920s and 1930s, frequently placed the largest individuals at shoulder heights of 5 to 5.5 metres and body masses as high as 30 tonnes; Granger and Gregory's 1936 reconstruction of Baluchitherium grangeri implied a shoulder height of approximately 5.25 metres.3 These early figures were clearly conservative in some directions and inflated in others, and they have been progressively revised downward as quantitative methods have improved.10, 15
The most influential modern reassessment is the volumetric body-mass analysis published by Asier Larramendi in 2016 in Acta Palaeontologica Polonica. Using the Graphic Double Integration method on detailed skeletal reconstructions, Larramendi estimated that the largest known individuals of P. transouralicum, represented by two cervical vertebrae and a metacarpal, corresponded to animals with shoulder heights of approximately 4.8 metres and body masses of about 17.1 tonnes (37,700 lb).7 Average adults are estimated at body masses closer to 11 tonnes, with the upper end of the range plausibly extending to 15–20 tonnes for the largest individuals; the older 30-tonne figures are now treated as unsupported.6, 7 Body length is generally reconstructed at approximately 7.4 metres from snout to tail base, following Vera Gromova's 1959 assessment, and head height while standing approached 6 metres.6
Even at the conservative Larramendi figures, Paraceratherium was several times the body mass of any extant terrestrial mammal: a large bull African elephant weighs roughly 6 tonnes, and the largest reliably weighed African elephant on record came in at about 10.4 tonnes. The only fossil mammal that may credibly have surpassed Paraceratherium in body mass is the Pleistocene straight-tusked elephant Palaeoloxodon namadicus, for which Larramendi has estimated body masses up to 22 tonnes from a fragmentary femur, though the underlying material is sufficiently incomplete that some authors continue to regard Paraceratherium as the largest known land mammal by complete skeletal remains.7
Body mass of Paraceratherium compared with other giant land mammals7, 15
Distribution and Eurasian biogeography
Remains assignable to Paraceratherium have now been recovered from early to late Oligocene formations across a broad swathe of Eurasia, including Pakistan, India, Kazakhstan, Mongolia, China, Georgia, Turkey, Romania, Bulgaria and adjacent parts of the Balkans. The geographic centre of diversity lies in Central and East Asia, with the Pakistani P. bugtiense representing the southern extension of the genus along the Tethyan margin and the Balkan and Anatolian occurrences marking its westernmost reach.6, 13 No Paraceratherium material has been recovered from the Americas, Africa or Australia, and the genus appears to have been entirely confined to the Northern Hemisphere of the Old World throughout its 11-million-year history.13
The 2021 description of P. linxiaense by Deng and colleagues substantially revised the picture of how the genus moved across Asia. Phylogenetic analysis of the new material recovered P. linxiaense from northwestern China and P. lepidum from Kazakhstan as sister taxa, with both nested next to P. bugtiense from Pakistan. The implication is that the lineage leading to P. bugtiense dispersed southward across the future Tibetan region and that the lineage leading to P. linxiaense subsequently retraced part of that route in the opposite direction during the late Oligocene.8 Such dispersals would have been impossible across a high Tibetan Plateau of the kind that exists today, and Deng and colleagues argued that they constitute strong palaeobiological evidence that much of the Tibetan region remained at relatively low elevation — perhaps below 2,000 metres — until well after the Oligocene-Miocene boundary, consistent with at least some palaeoaltimetric and clay-mineralogical reconstructions of regional uplift history.8, 14
The Hsanda Gol Formation of the Tsagan Nor Basin in central Mongolia, dating to the early Oligocene Rupelian stage approximately 31–32 million years ago, has produced some of the most informative Paraceratherium material outside of Pakistan and China. The formation preserves a transition from comparatively humid woodlands to a drier, more open landscape with ephemeral rivers and partially vegetated dune fields, and the associated mammalian fauna includes early rodents, lagomorphs and a range of smaller perissodactyls and artiodactyls in addition to giant indricotheres.12 The persistence of Paraceratherium across this aridification interval suggests that the genus was reasonably tolerant of habitat change and not narrowly tied to closed-canopy forest, an inference consistent with the broader Eurasian distribution of the group.12, 17
Ecology, feeding and life history
All available lines of evidence converge on the interpretation of Paraceratherium as a high-browsing herbivore that fed on the foliage of trees and shrubs in dry to semi-arid scrubland and open woodland environments. The simple, low-crowned cheek teeth are unsuited to processing tough, abrasive grasses and are appropriate for relatively soft leaves, twigs and shoots; carbon isotope analyses of tooth enamel from Asian specimens indicate a diet dominated by C3 plants, consistent with browsing in a world that pre-dated the late Miocene expansion of C4 grasslands.6, 17 The animal's height — with the head approaching 6 metres above the ground — would have allowed it to exploit canopy foliage well above the reach of any other contemporary herbivore, in a manner comparable to that of modern giraffes and okapis but on a vastly larger scale.6, 10
As a hindgut fermenter, Paraceratherium would have extracted relatively little energy per unit of plant material consumed and would consequently have needed to ingest enormous quantities of vegetation each day. Comparative scaling against modern hindgut fermenters such as horses and rhinos suggests daily food intake of several hundred kilograms of fresh leaves and browse, which in turn implies very large individual home ranges. Prothero estimates that an adult Paraceratherium would have required a home range on the order of 1,000 square kilometres or more to obtain sufficient browse, a figure consistent with allometric scaling of home-range area against body mass in extant megaherbivores.6 Population densities would correspondingly have been very low, and individuals were probably nomadic, perhaps moving in small family groups consisting of females with their dependent offspring.6
The combination of large body size, long generation time and low population density implies that Paraceratherium, like all megaherbivores, was a slow-reproducing K-strategist with limited demographic resilience. Comparable extant species such as elephants and rhinos have gestation periods of 16 to 22 months, single births, and inter-birth intervals of three to five years; an animal of Paraceratherium proportions would presumably have been at the upper end of this range, with calves taking many years to reach physical and reproductive maturity. These life-history constraints have featured prominently in subsequent discussions of why the genus eventually went extinct.6, 15
Paleoenvironment of the Oligocene Asian interior
The Oligocene world inhabited by Paraceratherium was strikingly different from the modern Asian interior. Global climate had cooled abruptly at the Eocene-Oligocene boundary approximately 34 million years ago, when the opening of the Drake Passage and the development of the Antarctic Circumpolar Current triggered the formation of the first permanent Antarctic ice sheets and a major drop in atmospheric carbon dioxide. The associated faunal turnover, known in Europe as the Grande Coupure and in Mongolia as the Mongolian Remodeling, replaced many archaic Eocene mammalian lineages with more modern groups, and it was in the immediate aftermath of this transition that the giant indricotheres rose to prominence.14, 19
In Asia, the Eocene-Oligocene transition was accompanied by a marked aridification of the continental interior, driven in part by the retreat of the Tethys Sea and the early stages of the Indo-Asian collision. Palaeosol and clay-mineralogical records from the northern Tibetan region indicate a shift from comparatively humid, forested conditions in the Eocene to more seasonal and drier conditions in the Oligocene, with mosaic landscapes of riparian woodland, open shrubland and ephemeral wetlands replacing the earlier closed forests.14 The Hsanda Gol Formation of Mongolia preserves precisely this kind of transitional environment, with vertebrate and small-mammal faunas indicating a fragmented woodland and shrubland landscape under a strongly seasonal climate.12
The Bugti beds of Pakistan, by contrast, preserve evidence of a wetter and more subtropical environment, with riparian forest and large mammals indicative of perennial water sources along the southern Tethyan margin. The Linxia Basin of northwestern China, which has produced both P. huangheense and P. linxiaense, was likewise relatively well-watered and supported a diverse Oligocene mammalian assemblage that included multiple species of giant rhino alongside chalicotheres, anthracotheres, and early proboscideans.8, 18 The collective picture is one of a genus that ranged across a broad latitudinal and ecological gradient within Asia, from comparatively humid southern environments to the seasonally arid Mongolian interior, and that was able to maintain very large body size across all of these settings.13, 17
Extinction at the Oligocene-Miocene boundary
Paraceratherium disappears from the Eurasian fossil record at or shortly before the Oligocene-Miocene boundary, approximately 23 million years ago, after a genus duration of roughly 11 million years. No definitive remains of the genus or its close relatives are known from any Miocene deposit, and the extinction appears to have been geologically abrupt, at least at the resolution of available stratigraphic sampling.6, 21 The disappearance of the giant rhinos forms part of a broader Oligocene-Miocene faunal turnover in Asia, in which several long-lived archaic groups were replaced by modern families that would dominate the Neogene.21
No single explanation for the extinction has gained consensus support, and Prothero argues in Rhinoceros Giants that monocausal hypotheses are unlikely to be correct given the scale and ecological breadth of the genus. Pure climate explanations are difficult to sustain because the genus had already weathered substantial Oligocene climatic and vegetational change without obvious decline; the body-size and life-history hypotheses likewise founder on the observation that Paraceratherium persisted for more than ten million years despite these constraints.6 The most plausible scenarios invoke the arrival of new competitors and predators as the principal driver. Beginning around 28 million years ago, gomphothere proboscideans dispersed northward out of Africa into Eurasia, introducing a new suite of large browsing herbivores that could plausibly have competed for canopy and sub-canopy foliage with Paraceratherium; over the same interval, a number of new carnivoran lineages arrived in Asia and may have increased predation pressure on calves and juveniles.6, 21
The combination of slow reproduction, very large home-range requirements and limited demographic flexibility characteristic of all extreme megaherbivores would have made Paraceratherium populations particularly vulnerable to even modest increases in adult or juvenile mortality from these new biotic factors, especially if reinforced by ongoing aridification and habitat fragmentation across the Asian interior. Ultimately, as Prothero notes, the true cause of the genus's disappearance will probably never be known with certainty, but the broad pattern is consistent with a slow ecological squeeze rather than a single catastrophic event.6, 15
Cultural and scientific legacy
Few extinct mammals have captured the public imagination as completely as Paraceratherium, in large part because the early twentieth-century reconstructions explicitly compared it to dinosaurs and presented it as a refutation of the popular assumption that mammals were inherently smaller than their Mesozoic predecessors. Henry Fairfield Osborn's 1923 announcement of Baluchitherium grangeri, the AMNH 18650 skull from Mongolia, attracted international press coverage and helped establish the giant rhinos as a recurring presence in popular books, museum exhibitions and natural-history documentaries throughout the twentieth century.16 The Granger and Gregory restoration of 1935 became the canonical visual depiction of the animal, reproduced in countless textbooks and posters, and remains the basis for the life-sized models displayed at the American Museum of Natural History and several other institutions today.4

{kind=link}
Within professional palaeontology, the genus has played an important role in calibrating the upper limits of terrestrial mammalian body size and in testing scaling models for megaherbivore biomechanics. Fortelius and Kappelman's 1993 paper, provocatively titled The largest land mammal ever imagined, used Paraceratherium as a reference point for examining the limits of column-supported limb design in graviportal mammals and the relationship between body mass and locomotor performance.10 Subsequent quantitative work by Smith and colleagues used Paraceratherium as a key data point in establishing the empirical maximum of mammalian body size and demonstrated that maximum body mass on each continent rose almost exponentially through the early Cenozoic before plateauing at values constrained by available land area and primary productivity.15
The continuing flow of new material — the 2017 description of P. huangheense, the 2021 announcement of P. linxiaense, and ongoing fieldwork in the Bugti Hills, the Linxia Basin and the Mongolian Gobi — demonstrates that the story of the giant rhinos is far from complete. Each new specimen has the potential to refine reconstructions of body mass, sharpen the phylogeny of Paraceratheriidae, and reshape the biogeographic models that link this extraordinary lineage to the broader history of mammalian evolution and the rise and fall of the Oligocene Asian fauna.8, 13, 18
References
Paraceratherium bugtiense, a new genus of Rhinocerotidae from the Bugti Hills of Baluchistan. Preliminary notice
On the skull and dentition of Paraceratherium bugtiense: a genus of aberrant rhinoceroses from the Lower Miocene deposits of Dera Bugti
Further notes on the gigantic extinct rhinoceros, Baluchitherium, from the Oligocene of Mongolia
A revised restoration of the skeleton of Baluchitherium, gigantic fossil rhinoceros of Central Asia
Earliest known unequivocal rhinocerotoid sheds new light on the origin of Giant Rhinos and phylogeny of early rhinocerotoids
New remains of the baluchithere Paraceratherium bugtiense (Pilgrim, 1910) from the late/latest Oligocene of the Bugti Hills, Balochistan, Pakistan
Tibetan plateau aridification linked to global cooling at the Eocene-Oligocene transition
Building Baluchitherium and Indricotherium: imperial and international networks in early-twentieth-century paleontology
Perissodactyl diversities and responses to climate changes as reflected by dental homogeneity during the Cenozoic in Asia
Postcranials of the giant rhino Paraceratherium huangheense (Mammalia, Perissodactyla) from the Early Oligocene Lanzhou Basin, NE Tibetan Plateau