Overview
- Plesiosaurs were a diverse order of marine reptiles (Plesiosauria) that thrived from the Early Jurassic to the end-Cretaceous extinction, spanning approximately 135 million years, and evolved two radically different body plans: the long-necked, small-headed plesiosauromorphs and the short-necked, large-headed pliosauromorphs, both propelled by a unique four-flipper locomotion system found in no other vertebrate group.
- Plesiosaur locomotion was powered by four large hydrofoil-shaped flippers in a flight-like underwater rowing motion, a biomechanically unique system among vertebrates that allowed precise manoeuvrability and sustained cruising, and their fossil record includes evidence of viviparity, deep diving, and a global distribution from tropical seas to polar waters.
- First scientifically described from specimens discovered by Mary Anning on the Jurassic coast of England in the 1820s, plesiosaurs were among the earliest fossil reptiles to challenge prevailing ideas about Earth history, and the entire order went extinct during the end-Cretaceous mass extinction 66 million years ago alongside non-avian dinosaurs, mosasaurs, and ammonites.
Plesiosaurs (order Plesiosauria, from the Greek plesios, "near to," and sauros, "lizard") were a major group of marine reptiles that dominated the world's oceans from the Early Jurassic to the end of the Cretaceous, a span of approximately 135 million years. Together with ichthyosaurs and mosasaurs, they constituted the three great lineages of Mesozoic marine reptiles, and their fossils are found on every continent, including Antarctica.1, 6 Plesiosaurs evolved a locomotion system unique among vertebrates — four large, hydrofoil-shaped flippers driven in a flight-like underwater rowing motion — and diversified into two radically different body plans that persisted throughout their evolutionary history: the long-necked, small-headed plesiosauromorphs and the short-necked, large-skulled pliosauromorphs.2
First scientifically described from specimens discovered by Mary Anning on the Jurassic coast of England in the 1820s, plesiosaurs were among the earliest large fossil reptiles to be recognised and played a foundational role in the development of vertebrate paleontology.15
{kind=link}
Origins and evolutionary relationships
Plesiosaurs belong to the larger clade Sauropterygia, which also includes the earlier Triassic marine reptiles known as nothosaurs and pachypleurosaurs. The sauropterygian lineage originated in the Early Triassic, shortly after the Permian-Triassic mass extinction, and underwent progressive adaptation to marine life over tens of millions of years. Nothosaurs, which ranged from the Early to Late Triassic, had partially webbed feet and probably alternated between aquatic and terrestrial locomotion, representing an intermediate stage in the transition from land to sea.6
Plesiosauria proper first appear in the fossil record in the latest Triassic (Rhaetian stage, approximately 205 million years ago), though they became diverse and abundant only in the Early Jurassic following the Triassic-Jurassic extinction at 201 million years ago. Phylogenetic analyses place plesiosaurs as the most derived sauropterygians, characterised by a fully rigid trunk, greatly enlarged and modified limb girdles, and four wing-like flippers adapted for continuous underwater propulsion.6, 10
Two body plans: plesiosauromorphs and pliosauromorphs
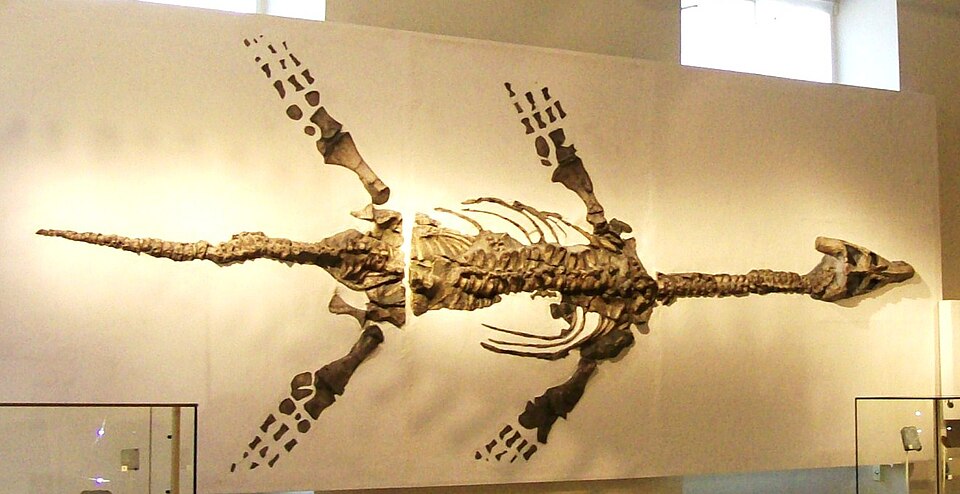
The most immediately striking feature of plesiosaur diversity is the repeated evolution of two contrasting body plans, traditionally designated as plesiosauromorph and pliosauromorph ecomorphotypes. Plesiosauromorphs had relatively small heads, extremely long necks containing up to 76 cervical vertebrae (more than any other known vertebrate), broad bodies, and four large flippers. Pliosauromorphs had proportionally enormous skulls — sometimes exceeding 2 metres in length — short, robust necks with fewer cervical vertebrae, and powerfully built bodies adapted for high-speed pursuit predation.2
O'Keefe demonstrated through quantitative analysis that these two body plans represent endpoints of a morphological continuum rather than two discrete clades. The plesiosauromorph body plan evolved independently in several plesiosaur families, including the Jurassic Plesiosauridae and Cryptoclididae and the Cretaceous Elasmosauridae, while the pliosauromorph body plan appeared in the Jurassic Pliosauridae and the Cretaceous Polycotylidae. This repeated convergence suggests strong ecological selection pressures favouring each body type for different predatory strategies.2
Plesiosaur body plan comparison2
| Feature | Plesiosauromorph | Pliosauromorph |
|---|---|---|
| Head size | Small relative to body | Very large (up to 2+ m) |
| Neck length | Long (up to 76 vertebrae) | Short (13–28 vertebrae) |
| Feeding strategy | Ambush / sweep feeding | Pursuit predation |
| Body length | 3–14 m | 4–12+ m |
| Example families | Elasmosauridae, Cryptoclididae | Pliosauridae, Polycotylidae |
Four-flipper locomotion
Plesiosaurs are the only known vertebrates to have employed four large flippers simultaneously as the primary means of propulsion. Unlike ichthyosaurs, which swam by oscillating the tail in a manner analogous to tuna and dolphins, plesiosaurs had relatively short, stiff tails that played little or no role in propulsion. Instead, all four limbs were modified into rigid, wing-shaped hydrofoils and moved in a subaqueous flight motion comparable to the wing beats of sea turtles or penguins, but with four propulsive surfaces rather than two.3
The biomechanics of four-flipper locomotion have been debated since the nineteenth century. Carpenter and colleagues used computational fluid dynamics and physical models to test competing hypotheses and concluded that plesiosaurs most likely employed a modified underwater flight stroke in which the fore and hind flippers operated asynchronously, with the forelimbs providing most of the thrust and the hindlimbs generating both thrust and stability. This system would have produced high manoeuvrability at moderate speeds rather than the sustained high-speed cruising achieved by the tail-driven ichthyosaurs, consistent with different predatory strategies.3
The anatomical basis for this locomotion system involved radical modification of the limb girdles. The pectoral and pelvic girdles of plesiosaurs were expanded into massive bony plates forming a ventral shield that anchored powerful flipper muscles. The limb bones themselves were shortened and flattened, and the digits were elongated through hyperphalangy (additional phalanges per digit) to create broad, rigid flippers with a high aspect ratio suited to generating lift in water.3, 10
The long neck and its function
The extreme elongation of the neck in plesiosauromorph plesiosaurs is one of the most remarkable anatomical features in the vertebrate fossil record. Elasmosaurid plesiosaurs of the Late Cretaceous possessed the most extreme necks, with Albertonectes vanderveldei holding the record at 76 cervical vertebrae and a neck exceeding 7 metres in length, representing more than half the total body length. The individual cervical vertebrae were relatively short and flat, and the overall neck structure was held together by interlocking articular surfaces and ligaments that would have restricted lateral flexibility while allowing ventral and dorsal bending.7, 12
The function of these extraordinarily long necks has been extensively debated. Early hypotheses suggested that long-necked plesiosaurs held their heads high above the water surface like swans, but biomechanical analysis has demonstrated that the neck could not have been raised far above the water surface without exceeding the structural limits of the cervical joints. Current consensus favours the hypothesis that the long neck was used primarily underwater, allowing the small head to approach schooling fish or cephalopods from below or from the side without the large body alerting prey. The neck may have functioned as a mobile hunting tool, sweeping through shoals of fish while the body remained relatively stationary.2, 12
Diet and ecology
Plesiosaurs occupied a wide range of ecological niches during their 135-million-year history. Direct evidence of diet comes from fossilised stomach contents, which have been preserved in several exceptional specimens. McHenry and colleagues described three-dimensionally preserved stomach contents from a Cretaceous plesiosaur in Queensland, Australia, containing the remains of fish, cephalopod beaks, and crustaceans, confirming a generalist marine predatory diet for at least some species.13
Plesiosauromorph taxa with small heads and long necks were likely ambush or sweep feeders, using their necks to approach prey stealthily while the body remained at a distance. Their teeth were typically long, slender, and interlocking, forming a trap-like array suited for grasping slippery fish and squid. Pliosauromorph taxa, by contrast, were pursuit predators with massive skulls, powerful jaw muscles, and robust conical teeth capable of subduing large prey. The largest pliosaurids, such as Pliosaurus and Liopleurodon, occupied apex predator niches and may have fed on other marine reptiles, large fish, and cephalopods.4, 9
Some plesiosaurs appear to have adopted specialised feeding strategies. Gastroliths (stomach stones) have been found associated with several plesiosaur specimens, though their function remains debated: proposed roles include ballast for buoyancy control, grinding stones for food processing, or incidentally swallowed material.13
Reproduction and viviparity
A landmark discovery in 2011 provided the first direct evidence that plesiosaurs were viviparous — they gave birth to live young rather than laying eggs on land. O'Keefe and Chiappe described a specimen of the Late Cretaceous polycotylid Polycotylus latippinus from Kansas containing a single large foetus within the body cavity. The foetus was approximately one-third the length of the mother, indicating that plesiosaurs produced relatively large, precocial offspring in small litter sizes — a K-selected reproductive strategy comparable to that of modern marine mammals such as whales and dolphins.5
The discovery of viviparity resolved a longstanding question about plesiosaur biology. Unlike sea turtles, which must return to land to lay eggs, plesiosaurs with their rigidly adapted flippers and massive body proportions could not have hauled themselves onto beaches. Viviparity freed them from any dependence on terrestrial environments and allowed fully oceanic lifestyles. O'Keefe and Chiappe further suggested that the small litter size and large offspring size implied extended parental care, possibly involving group social behaviours analogous to those of modern cetaceans, though direct evidence for such behaviours is lacking.5
Geographic distribution
Plesiosaur fossils have been found on every continent, including Antarctica, demonstrating a truly global distribution. During the Jurassic and Cretaceous, when sea levels were significantly higher than today and many modern landmasses were submerged or connected by shallow seaways, plesiosaurs inhabited a wide range of marine environments from tropical to polar waters.8, 11
Particularly significant are the discoveries of plesiosaurs from high-latitude sites. Elasmosaurid plesiosaurs from the Maastrichtian (latest Cretaceous) of Antarctica, including specimens from James Ross Island and Seymour Island, demonstrate that these animals thrived in polar waters where seasonal sea ice may have been present. O'Gorman and colleagues described a palaeopolar elasmosaurid from the Maastrichtian of Antarctica that represents one of the last plesiosaurs before the end-Cretaceous extinction, extending the group's temporal range to within the final few hundred thousand years before the impact.8, 11
Plesiosaurs also penetrated freshwater environments. Several species have been found in non-marine deposits, including river and lake sediments, suggesting that at least some plesiosaurs were capable of tolerating reduced salinity, much as modern bull sharks move between saltwater and freshwater habitats.1
Discovery and scientific history
The scientific recognition of plesiosaurs began in the early nineteenth century on the Jurassic coast of southern England. Mary Anning (1799–1847), the pioneering fossil collector from Lyme Regis, discovered the first nearly complete plesiosaur skeleton in December 1823, following her earlier discovery of an ichthyosaur skeleton in 1811. The plesiosaur specimen, which Anning excavated from the Blue Lias cliffs, was described by William Conybeare in 1824 as Plesiosaurus dolichodeirus and caused a sensation in the scientific world because of its extraordinary body plan — the combination of a long neck, small head, broad body, and four large flippers was unlike any living animal.15, 14
The anatomist Georges Cuvier initially expressed scepticism about the specimen, suspecting it might be a forgery, but examination of the original bones confirmed their authenticity. Richard Owen later included plesiosaurs in his comprehensive surveys of fossil reptiles and helped establish their position within reptilian classification. Throughout the nineteenth century, plesiosaur discoveries expanded to include specimens from Germany, France, Australia, and North America, revealing the group's global distribution and morphological diversity.14, 10
Extinction at the K-Pg boundary
Unlike ichthyosaurs, which went extinct approximately 94 million years ago during the Cenomanian-Turonian oceanic anoxic event, plesiosaurs survived until the very end of the Cretaceous and were exterminated by the end-Cretaceous mass extinction at 66 million years ago. This event, triggered by the Chicxulub asteroid impact and possibly exacerbated by Deccan Traps volcanism, eliminated approximately 76 percent of all species on Earth, including all non-avian dinosaurs, mosasaurs, ammonites, and the remaining plesiosaurs.16
Benson and Druckenmiller analysed plesiosaur diversity patterns through the Cretaceous and found that the group maintained moderate diversity and ecological disparity right up to the K-Pg boundary, with no evidence of a prolonged decline preceding the extinction. Both elasmosaurid plesiosauromorphs and polycotylid pliosauromorphs were present in the latest Maastrichtian, suggesting that the plesiosaur extinction was an abrupt event caused by the environmental catastrophe of the impact rather than the culmination of a gradual decline. This pattern contrasts markedly with the ichthyosaurs, whose extinction was drawn out over millions of years, and indicates that plesiosaurs as a group were ecologically viable until external catastrophic forcing eliminated them.16
References
Viviparity and K-selected life history in a Mesozoic marine plesiosaur (Reptilia, Sauropterygia)
Aristonectes quiriquinensis, sp. nov., a new highly derived elasmosaurid from the upper Maastrichtian of central Chile
A palaeopolar elasmosaurid (Sauropterygia; Plesiosauria) from the Maastrichtian of Antarctica
A new polycotylid plesiosaur with a unique dentition from the Upper Cretaceous of the Williston Basin
Plesiosauria from the Upper Cretaceous (Campanian) of the James Ross Basin, Antarctica