Overview
- Velociraptor was a small, feathered dromaeosaurid theropod that lived approximately 75 to 71 million years ago in what is now Mongolia and China, reaching only about 2 metres in length and 15 to 20 kilograms in body mass — far smaller than its depiction in popular media.
- The famous ‘fighting dinosaurs’ specimen, discovered in 1971, preserves a Velociraptor locked in combat with a Protoceratops, providing one of the most dramatic and direct pieces of evidence for predatory behaviour in non-avian dinosaurs.
- Phylogenetic analyses place Velociraptor within Dromaeosauridae, a family of maniraptoran theropods closely related to birds, and quill knob evidence from the closely related Velociraptor mongoliensis confirms the presence of well-developed pennaceous feathers on its forelimbs.
Velociraptor is a genus of small dromaeosaurid theropod dinosaur that lived during the Late Cretaceous period, approximately 75 to 71 million years ago, in what is now Mongolia and northern China. First described by Henry Fairfield Osborn in 1924 from fossils collected during the American Museum of Natural History's Central Asiatic Expeditions, Velociraptor has become one of the most scientifically important non-avian dinosaurs for understanding the evolutionary transition from non-avian theropods to birds.1, 10 The type species, Velociraptor mongoliensis, is known from numerous specimens recovered from the Djadochta Formation of the Gobi Desert, while a second species, V. osmolskae, has been described from the Bayan Mandahu Formation of Inner Mongolia, China.1, 15
{kind=link}
Despite its enormous fame in popular culture — driven primarily by its dramatically enlarged and inaccurate portrayal in the 1993 film Jurassic Park — the real Velociraptor was a modestly sized animal, approximately 2 metres in total length and weighing an estimated 15 to 20 kilograms, roughly comparable in size to a large turkey. Its scientific significance lies not in its size but in the exceptional quality of its fossil record, which includes one of the most famous specimens in all of palaeontology: the "fighting dinosaurs," a Velociraptor preserved in mortal combat with a Protoceratops.11, 17
Discovery and naming
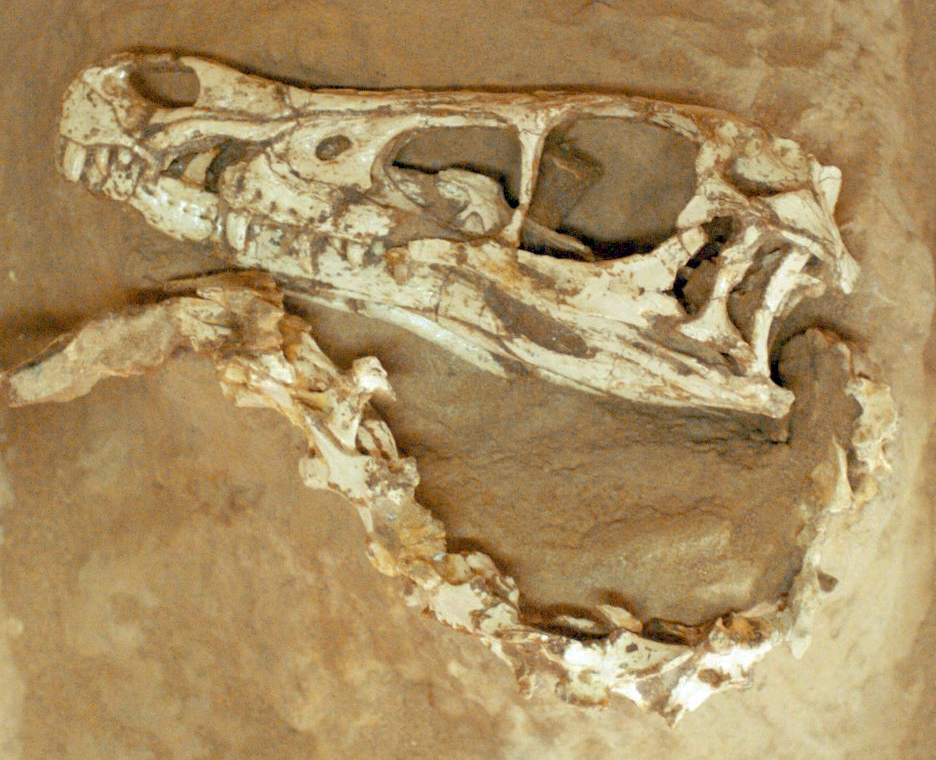
The first specimen of Velociraptor was discovered on 11 August 1923 by Peter Kaisen, a member of the American Museum of Natural History's Central Asiatic Expeditions led by Roy Chapman Andrews, at the Flaming Cliffs locality (Bayan Zag) in the Djadochta Formation of the Ömnögovi Province, southern Mongolia. The specimen (AMNH 6515) consisted of a nearly complete but crushed skull approximately 23 centimetres in length, along with the distinctive sickle-shaped second pedal ungual that would become one of the animal's most recognisable features. Henry Fairfield Osborn described and named the genus in 1924, choosing the name Velociraptor ("swift seizer") in reference to the animal's presumed speed and predatory habits, and erecting the species V. mongoliensis to indicate its Mongolian provenance.1
Following the cessation of American expeditions to Mongolia after 1930, fieldwork in the Gobi Desert was carried out primarily by Soviet-Mongolian and Polish-Mongolian expeditions in the 1960s and 1970s. These expeditions recovered numerous additional specimens of Velociraptor from the Djadochta Formation, substantially expanding the anatomical record of the genus. Among the most significant finds was a specimen described by Barsbold in 1974, which added important postcranial information.2 Barsbold and Osmólska's 1999 study provided a detailed redescription of the skull anatomy based on multiple specimens, clarifying features of the lacrimal, postorbital, and quadrate that had been obscured in the crushed holotype.12
A second species, Velociraptor osmolskae, was described in 2008 by Godefroit and colleagues on the basis of a partial skull (IMM 99NM-BYM-3/3) from the Bayan Mandahu Formation of Inner Mongolia, China. This formation is considered approximately contemporaneous with the Djadochta Formation and shares much of its fauna, including Protoceratops and oviraptorid dinosaurs. V. osmolskae differs from V. mongoliensis primarily in features of the maxilla and the frontal-lacrimal contact, and its recognition extended the geographic range of the genus from Mongolia into northern China.15
Anatomy and body plan
Velociraptor was a lightly built, bipedal predator with a body plan broadly typical of dromaeosaurid theropods. The skull was long and low, measuring approximately 23 to 25 centimetres in length, with an elongate, dorsoventrally flattened rostrum that distinguishes it from the deeper, shorter skulls of many other dromaeosaurids such as Dromaeosaurus and Deinonychus. The premaxilla bore four teeth on each side, the maxilla approximately eleven, and the dentary some fifteen, giving a total tooth count of approximately sixty. The teeth were serrated along both anterior and posterior carinae, moderately recurved, and widely spaced — a morphology consistent with a predator that seized and held relatively small prey rather than slicing through large carcasses.7, 12
The postcranial skeleton of Velociraptor reflects an animal adapted for agility rather than brute force. The vertebral column comprised approximately ten cervical vertebrae forming a sinuous, S-shaped curve, thirteen dorsals, six sacrals fused into a solid synsacrum, and approximately thirty caudal vertebrae. The tail was stiffened along most of its length by elongate prezygapophyses and chevron processes that extended anteriorly and posteriorly over several adjacent vertebrae, effectively transforming the distal tail into a rigid rod. This construction would have provided dynamic balance during rapid turns and leaps, functioning as a counterbalance to the body's anterior mass during swift manoeuvres.10, 17
The forelimbs were relatively long and bore three-fingered hands with strongly recurved claws. The wrist contained a semilunate carpal block that permitted the hand to fold against the forearm in a motion homologous with the wing-folding mechanism of modern birds. This wrist structure, shared across Maniraptora, is one of numerous skeletal features linking dromaeosaurids to the avian lineage.10 The hindlimbs were proportionally long relative to body size, with the tibia exceeding the femur in length — a limb proportion associated with cursoriality in theropods. The metatarsus was elongate, and the foot bore three functional weight-bearing toes (digits II, III, and IV), with digit I reduced and positioned higher on the metatarsus.
The most celebrated anatomical feature of Velociraptor is the enlarged, sickle-shaped ungual claw on the second digit of each foot. This claw was strongly recurved, laterally compressed, and held in a retracted, hyperextended position during locomotion, keeping its sharp point clear of the ground. In V. mongoliensis, the second pedal ungual measured approximately 6.5 centimetres along the outer curve. The functional interpretation of this claw has been debated, with hypotheses ranging from disembowelling prey to climbing and pinning. Biomechanical analyses of claw geometry in dromaeosaurids suggest that the claw was better suited for piercing and hooking than for slashing, and the "fighting dinosaurs" specimen demonstrates its deployment in a predatory context.7, 10
Feathered integument
The question of whether Velociraptor bore feathers was resolved definitively in 2007 when Turner, Makovicky, and Norell reported the presence of quill knobs on the ulna of a V. mongoliensis specimen (IGM 100/981) from the Djadochta Formation. Quill knobs are small, regularly spaced bony bumps on the posterior surface of the ulna that serve as attachment points for the ligaments anchoring large secondary flight feathers (or pennaceous feathers homologous with secondaries) in modern birds. Their presence in Velociraptor constitutes unambiguous osteological evidence that the animal possessed well-developed feathers along its forelimbs, despite the absence of direct feather impressions in Djadochta Formation specimens, which are preserved in aeolian sandstone rather than the fine-grained lacustrine sediments that typically preserve integumentary structures.8
The discovery of quill knobs in Velociraptor did not come as a surprise to palaeontologists, given the abundant evidence for feathered integument across Maniraptora by 2007. The closely related dromaeosaurid Sinornithosaurus, described from the Early Cretaceous Yixian Formation of Liaoning Province, China, was found preserved with filamentous integumentary structures interpreted as protofeathers or Stage I feathers.14 Microraptor, another dromaeosaurid from the same deposits, possessed fully developed asymmetric flight feathers on both its forelimbs and hindlimbs, demonstrating that powered or gliding flight existed within Dromaeosauridae before the divergence of birds.6 The phylogenetic bracket established by these feathered forms, combined with the direct osteological evidence from quill knobs, makes it virtually certain that Velociraptor was covered in a coat of feathers over much of its body, though these feathers would not have enabled flight in an animal of its size and skeletal proportions.
The presence of well-developed pennaceous feathers in a flightless dromaeosaurid raises important questions about the original function of feathers in the dinosaur-bird transition. Turner and colleagues noted that quill knobs are most prominent in birds that use their feathers for aerodynamic purposes, suggesting that the feathered forelimbs of Velociraptor may have been inherited from smaller, volant ancestors and subsequently retained for functions such as display, brooding, or stabilisation during prey capture, even as the lineage evolved body sizes incompatible with flight.8, 18
The fighting dinosaurs
The single most famous Velociraptor specimen is GIN 100/25, the "fighting dinosaurs," discovered in 1971 by a Polish-Mongolian palaeontological expedition at the Tugriken Shireh (Tögrögiin Shiree) locality in the Djadochta Formation of southern Mongolia. The specimen preserves a Velociraptor mongoliensis and a Protoceratops andrewsi locked together in what appears to be mortal combat, entombed simultaneously by a catastrophic burial event — most likely the rapid collapse of a sand dune or the sudden onset of a sandstorm that buried both animals alive before they could separate.11
The taphonomic details of the specimen are extraordinarily informative. The Velociraptor is positioned on its right side with its left foot extended toward the throat region of the Protoceratops, the sickle claw embedded in the area corresponding to the carotid artery or jugular vein. This posture strongly suggests that the claw was being used as a puncturing weapon directed at the vulnerable ventral neck of the prey, consistent with the hypothesis that dromaeosaurid sickle claws functioned primarily as piercing implements rather than slashing tools. The Protoceratops, for its part, has the right forelimb of the Velociraptor clamped firmly in its beak, and the Velociraptor's arm appears to be broken at the point of contact, suggesting that the ceratopsian's powerful beak inflicted serious damage on the predator even as it was being attacked.11, 7
The fighting dinosaurs specimen provides some of the most direct fossil evidence for predator-prey interactions among non-avian dinosaurs. The posture of the two animals has been interpreted as capturing the moment of a predatory attack rather than scavenging behaviour, as the positions of the limbs and the active engagement of both animals indicate ongoing struggle at the time of burial. The specimen has been designated a national treasure of Mongolia and is housed at the Mongolian Academy of Sciences in Ulaanbaatar, though casts are exhibited in museums worldwide.11
Hunting behaviour and diet
The predatory ecology of Velociraptor has been interpreted primarily on the basis of its dental and claw morphology, the fighting dinosaurs specimen, and comparisons with other dromaeosaurids for which more extensive taphonomic data are available. The relatively small body size of Velociraptor — approximately 15 to 20 kilograms — constrains the range of prey it could have tackled as a solitary predator. Adult Protoceratops, which reached body masses of approximately 80 to 180 kilograms, would have represented large and dangerous prey for a lone Velociraptor, and the fighting dinosaurs specimen demonstrates that such encounters could be lethal for the predator as well as the prey.7, 17
The question of whether dromaeosaurids hunted in coordinated packs, as depicted in popular media, remains contentious. The hypothesis of pack hunting in Velociraptor is based primarily on analogy with the larger dromaeosaurid Deinonychus, for which multiple individuals have been found associated with the herbivore Tenontosaurus at several localities in the Cloverly Formation of Montana and Wyoming. However, Roach and Brinkman argued in 2007 that these associations more closely resemble the feeding behaviour of extant Komodo dragons, in which multiple individuals converge opportunistically on a single carcass, than the coordinated social hunting of wolf packs. No definitive evidence for cooperative hunting has been demonstrated in Velociraptor specifically, and the absence of multi-individual bonebeds attributable to this genus argues against routine pack behaviour.3, 17
The sensory capabilities of Velociraptor provide additional insight into its predatory ecology. CT scanning of the braincase has revealed that Velociraptor possessed a relatively large brain for its body size, with an encephalisation quotient at the upper end of the non-avian dinosaur range. The olfactory bulbs were moderately developed, the optic lobes were large, and the floccular lobes of the cerebellum — associated with gaze stabilisation and coordination of rapid head and eye movements — were proportionally expanded, suggesting an animal that relied heavily on vision during rapid, agile predatory manoeuvres. The semicircular canals of the inner ear indicate sensitivity to rapid rotational accelerations of the head, consistent with an active, visually guided predator rather than a scavenger or ambush hunter.10, 17
The diet of Velociraptor likely encompassed a range of small to medium-sized vertebrates available in its semi-arid environment. The Djadochta Formation fauna includes abundant Protoceratops of various ontogenetic stages, small ornithischians, lizards, mammals, and other small theropods. Juvenile Protoceratops, much smaller and less well defended than adults, may have formed a significant component of the diet. The serrated, widely spaced teeth of Velociraptor are consistent with a generalist predatory diet rather than specialisation on any single prey type, and the animal's speed and agility would have been well suited to pursuing small, fast-moving prey across the dune fields and interdune flats of the Djadochta palaeoenvironment.7, 13
Dromaeosaurid relationships and the bird connection
Velociraptor is classified within the family Dromaeosauridae, a clade of maniraptoran theropod dinosaurs that includes some of the closest known non-avian relatives of birds. Within Dromaeosauridae, phylogenetic analyses consistently place Velociraptor in the subfamily Velociraptorinae, which also includes Tsaagan (from the same Djadochta Formation), Linheraptor, and possibly Deinonychus, though the position of the latter taxon varies among analyses. The most comprehensive phylogenetic treatment of the family is Turner, Makovicky, and Norell's 2012 monograph, which scored 111 taxa across 474 characters and recovered a well-resolved topology in which Dromaeosauridae comprises several major subclades: Velociraptorinae, Dromaeosaurinae, Microraptorinae, and the enigmatic Unenlagiinae of the Southern Hemisphere.10
The relationship between dromaeosaurids and birds is central to one of the most transformative findings in modern evolutionary biology. Dromaeosauridae is nested within Maniraptora, a clade that also includes Troodontidae and Avialae (the group containing Archaeopteryx and all modern birds). Together, Dromaeosauridae and Troodontidae form the clade Deinonychosauria, which is recovered as the sister group of Avialae in most phylogenetic analyses, making dromaeosaurids among the closest relatives of birds outside Avialae itself.9, 10 This phylogenetic position means that Velociraptor and modern birds shared a common ancestor more recently than either did with other major theropod groups such as tyrannosaurids or ornithomimids.
The shared features between dromaeosaurids and birds are extensive and include not only feathered integument and the semilunate carpal block in the wrist, but also a furcula (wishbone), hollow pneumatic bones, a posteriorly directed pubis, and numerous details of skull, vertebral, and pelvic anatomy. Lee and colleagues' 2014 analysis of body mass evolution across theropods demonstrated that the lineage leading to birds experienced sustained miniaturisation over approximately 50 million years, with dromaeosaurids representing a branch that reversed this trend, secondarily evolving larger body sizes after diverging from the common ancestor shared with birds.18 This finding suggests that Velociraptor is not an evolutionary "step" toward birds but rather a cousin that independently evolved in its own direction after the split from the avian line.
Comparative anatomy of Velociraptor and selected dromaeosaurids10, 17
| Taxon | Body length (m) | Mass (kg) | Age (Ma) | Region |
|---|---|---|---|---|
| Velociraptor mongoliensis | ~2.0 | 15–20 | 75–71 | Mongolia |
| Deinonychus antirrhopus | ~3.4 | 70–100 | 115–108 | North America |
| Microraptor zhaoianus | ~0.8 | 1–2 | 125–120 | China |
| Utahraptor ostrommaysi | ~5.0 | 300–500 | 139–134 | North America |
| Dromaeosaurus albertensis | ~2.0 | 15–20 | 77–74 | North America |
Palaeoecology of the Djadochta Formation
The Djadochta Formation, from which the majority of Velociraptor specimens have been recovered, records a Late Cretaceous (Campanian, approximately 75 to 71 Ma) semi-arid to arid continental environment in what is now the southern Gobi Desert of Mongolia. The formation is characterised by extensive cross-bedded aeolian sandstones representing migrating dune fields, interbedded with finer-grained interdune and fluvial deposits. The sedimentology indicates a landscape dominated by sand dunes with ephemeral oases, intermittent streams, and vegetated interdune flats, analogous in some respects to modern semi-arid environments such as the margins of the Namib or Gobi deserts.13
The fauna of the Djadochta Formation is remarkably well known, thanks in part to the exceptional preservational conditions afforded by rapid burial in wind-blown sand. In addition to Velociraptor, the formation has yielded the ceratopsian Protoceratops andrewsi (by far the most abundant dinosaur in the assemblage), the oviraptorid Oviraptor philoceratops, the troodontid Saurornithoides mongoliensis, the ankylosaur Pinacosaurus grangeri, and a diverse assemblage of lizards and mammals including multituberculates and early placental mammals. The high proportion of small-bodied vertebrates and the relative absence of large-bodied herbivores and predators distinguish the Djadochta fauna from contemporaneous formations in North America such as the Dinosaur Park Formation, which was deposited in more mesic, coastal-plain environments and supported a far greater diversity of large-bodied dinosaurs.13, 17
The taphonomic conditions of the Djadochta Formation have profoundly shaped what is known about Velociraptor and its contemporaries. Rapid burial by collapsing dunes or sandstorms accounts for the articulated preservation of many specimens, including the fighting dinosaurs, nesting oviraptorids, and curled-up Protoceratops. These catastrophic burial events captured snapshots of animal behaviour — predation, nesting, sheltering — that are extraordinarily rare in the fossil record and have provided some of the most vivid windows into the daily lives of non-avian dinosaurs.11, 13
Popular culture versus scientific reality
No dinosaur genus exhibits a greater disparity between its scientific reality and its popular image than Velociraptor. The portrayal of Velociraptor in the 1993 film Jurassic Park and its sequels — as a human-sized, scaly, hyperintelligent pack hunter capable of opening door handles — bears little resemblance to the actual animal. The film's velociraptors were modelled primarily on Deinonychus antirrhopus, a substantially larger dromaeosaurid from the Early Cretaceous of North America, and the decision to use the name Velociraptor instead was reportedly influenced by the fact that author Michael Crichton considered it more dramatic. The film's animals are depicted at approximately 1.8 metres tall at the hip and over 3 metres in length, whereas the real Velociraptor mongoliensis stood only about 0.5 metres at the hip and was approximately 2 metres long including its tail.17
The absence of feathers in the film's depiction is another significant departure from scientific understanding. Although direct evidence for feathered integument in Velociraptor was not published until 2007, the phylogenetic position of dromaeosaurids within Maniraptora meant that feathered integument was predicted for Velociraptor long before the quill knob discovery confirmed it. By the time the discovery was made, the feathered dromaeosaurid Sinornithosaurus had already been known for six years. The continued depiction of scaly velociraptors in subsequent Jurassic Park sequels has been a source of frustration for palaeontologists, though the franchise's cultural influence has also generated enormous public interest in dinosaur science.8, 14
The question of intelligence is similarly exaggerated in popular culture. While dromaeosaurids possessed relatively large brains for their body size compared to most non-avian dinosaurs, and their encephalisation quotients are toward the upper end of the non-avian dinosaur range, there is no evidence that Velociraptor possessed cognitive abilities remotely comparable to those attributed to it in fiction. Its brain was comparable in relative size to those of modern ratite birds such as ostriches and emus, suggesting competent predatory behaviour and reasonably complex sensory processing, but nothing approaching the problem-solving capabilities depicted on screen.17
The cultural impact of Jurassic Park on public perception of Velociraptor has created both opportunities and challenges for science communication. On one hand, the film generated a wave of public interest in dinosaurs that persists to the present day and has contributed to increased funding for palaeontological research and museum attendance. On the other hand, the deeply embedded popular image of Velociraptor as a large, scaly, pack-hunting predator requires constant correction, and many popular accounts of the animal continue to conflate it with Deinonychus or reproduce the film's inaccuracies without qualification. The real Velociraptor — a small, feathered, solitary or loosely social predator of the Mongolian dune fields — is in many ways a more remarkable animal than its cinematic counterpart, representing as it does a window into the evolutionary radiation that ultimately produced the 10,000-plus species of living birds.8, 18
References
A new Velociraptorinae (Dinosauria: Dromaeosauridae) from the Upper Cretaceous of northern China
A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus
New specimen of Velociraptor mongoliensis (Theropoda: Dromaeosauridae) from the Djadokhta Formation, and the use of didactyl tracks
The fighting dinosaurs: the position of a remarkable new find from the Cretaceous of Mongolia
The Djadokhta Formation correlatives in Chinese Inner Mongolia: an overview of the stratigraphy, sedimentary geology, and paleontology and comparisons with the type locality in the pre-Altai Gobi
A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China
A second specimen of Velociraptor osmolskae (Theropoda: Dromaeosauridae) from Inner Mongolia, China
Rates of dinosaur body mass evolution indicate 170 million years of sustained ecological innovation on the avian stem lineage