Overview
- The evolutionary emergence of language left no direct fossil record, forcing researchers to reconstruct its origins from indirect evidence including vocal tract anatomy, neurogenetics, archaeological proxies for symbolic cognition, and comparative studies of primate communication.
- The discovery that mutations in the FOXP2 gene cause severe speech and language disorders in the KE family, combined with evidence that the derived human variant was shared with Neanderthals, has made FOXP2 the best-studied gene in language evolution, though language clearly depends on many genes acting in concert.
- Whether language emerged gradually over hundreds of thousands of years through gestural, vocal, or musical protolanguage stages, or appeared rapidly as a consequence of a late neural reorganization, remains one of the most contested questions in the human sciences, with current evidence increasingly favoring a deep, incremental origin.
The origin of human language is one of the most difficult problems in science. Language does not fossilize: there are no preserved utterances, no imprints of syntax in stone, no direct physical traces of the moment our ancestors first used words to describe the world. Unlike bipedalism, which leaves its signature in fossilized pelves and footprints, or tool use, which is documented by millions of stone artefacts, language must be reconstructed entirely from indirect evidence — the shape of the vocal tract, the size of neural pathways, the genetics of speech-related disorders, and the archaeological proxies for symbolic thought that language presumably accompanied.2, 25 The question of how, when, and why language evolved has attracted contributions from linguistics, neuroscience, genetics, archaeology, and evolutionary biology, yet after more than a century of formal inquiry, no consensus has emerged on even the most basic issues: whether language arose gradually or suddenly, whether it began as gesture or vocalization, and whether its core properties are unique to humans or shared in rudimentary form with other species.1, 2
What is clear is that fully modern language is a species-universal trait of Homo sapiens. Every known human society possesses a richly structured language with phonology, morphology, syntax, and semantics, and all neurologically typical children acquire their native language without formal instruction within the first few years of life.1 No non-human animal has been shown to possess anything comparable in complexity, productivity, or displacement — the ability to refer to things absent in space and time. The challenge is to explain how this remarkable capacity arose in the hominin lineage, and the following sections survey the principal lines of evidence and the major theoretical frameworks that have been brought to bear on the problem.
Why language origins are difficult to study
The study of language origins faces a set of methodological obstacles that are unique among questions in human evolution. The most fundamental is the absence of direct evidence. Spoken language leaves no fossil trace, and the earliest writing systems appear only around 5,000 years ago — a vanishingly recent moment relative to the hundreds of thousands of years over which language likely evolved.2 Unlike bones, teeth, and stone tools, which preserve across geological time, the acoustic signals of speech and the cognitive operations that generate sentences are ephemeral. Researchers must therefore rely on indirect proxies: the anatomy of the vocal tract and the brain, the genetics of language-related disorders, the comparative study of communication in other species, and the archaeological record of behaviours that are thought to depend on or co-occur with language, such as symbolic art, long-distance trade, and complex technology.2, 25
A second difficulty is that language is not a single trait but a composite of many capacities — a sensory-motor system for producing and perceiving signals, a conceptual-intentional system for constructing meanings, and a computational system for combining units into hierarchically structured sequences.1 These components may have different evolutionary histories, different genetic underpinnings, and different degrees of continuity with the communication systems of other animals. The question "when did language evolve?" may therefore have no single answer; different aspects of language may have emerged at different times and under different selective pressures.2, 25
A third challenge is the comparative method itself. The closest living relatives of humans, chimpanzees and bonobos, diverged from the human lineage approximately six to seven million years ago, and all intermediate hominin species are extinct. It is therefore impossible to study the communication systems of our direct ancestors, and comparisons with extant apes reveal only what is shared with a very distant common ancestor or what has evolved convergently.1, 2 For these reasons, the Linguistic Society of Paris famously banned papers on language origins in 1866, and the topic was widely regarded as intractable until the late twentieth century, when advances in genetics, neuroimaging, and palaeoanthropology reopened the field on firmer empirical ground.25
Anatomical prerequisites for speech
{kind=link}
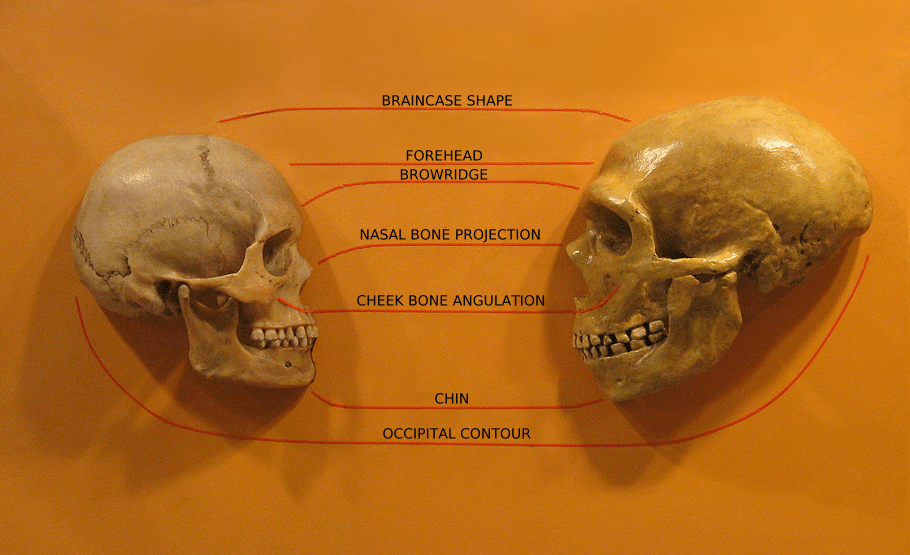
Speech production in modern humans depends on a suite of anatomical features that differ from those of other primates. The most widely discussed is the position of the larynx. In most mammals and in human infants, the larynx sits high in the throat, allowing the epiglottis to interlock with the soft palate and create separate channels for breathing and swallowing. In adult humans, the larynx descends to a lower position, creating an expanded pharyngeal cavity above the vocal folds that serves as a resonating chamber crucial for producing the full range of vowel sounds, particularly the "point vowels" /i/, /a/, and /u/ that anchor the vowel systems of all human languages.3 Lieberman and Crelin argued in 1971 that Neanderthals lacked a fully descended larynx and would therefore have been unable to produce these vowels, though this conclusion has been challenged by subsequent research showing that a descended larynx is not uniquely human — red deer and several other mammals lower the larynx during vocalisation — and that the relationship between laryngeal position and speech capability is more complex than initially proposed.3, 4
Another anatomical proxy is the hypoglossal canal, the bony channel in the base of the skull through which the hypoglossal nerve passes to innervate the muscles of the tongue. Kay, Cartmill, and Balow proposed in 1998 that the size of the hypoglossal canal reflects the degree of neural control over tongue movements, and reported that modern humans and Neanderthals have significantly larger canals than australopithecines and chimpanzees, suggesting that fine motor control of the tongue — essential for articulate speech — was present in the genus Homo by at least 400,000 years ago.5 However, DeGusta and colleagues contested this interpretation in 1999, demonstrating that several non-human primate species and early hominins also fall within the modern human range of hypoglossal canal size, which may therefore be a poor indicator of speech ability.6
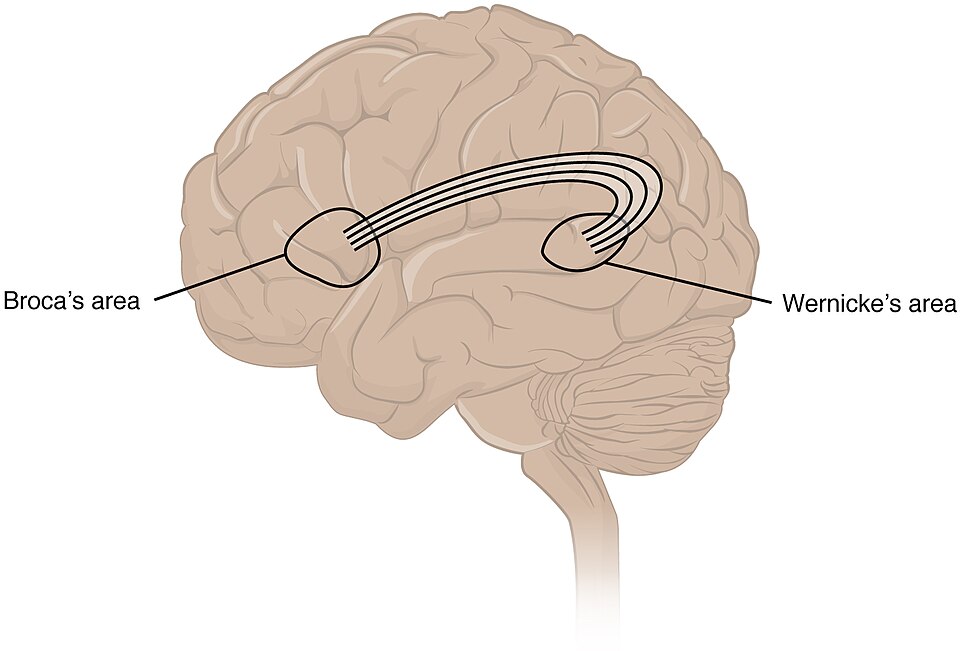
The neuroanatomy of language centres in the brain provides additional evidence. In modern humans, language processing depends critically on Broca's area in the left inferior frontal gyrus, which is involved in speech production, syntactic processing, and the hierarchical sequencing of motor actions, and Wernicke's area in the left posterior superior temporal gyrus, which is central to speech comprehension and semantic processing. These regions are connected by the arcuate fasciculus, a white-matter tract whose left-hemisphere dominance is a hallmark of the human brain.10 Endocranial casts of fossil hominins have been used to infer the presence of Broca's area as early as Homo habilis, approximately 1.8 million years ago, though whether its presence implies language-like function or merely complex motor planning remains debated.2
The FOXP2 gene and the genetics of speech
The genetics of language received a dramatic boost with the study of the KE family, a large three-generation British pedigree in which approximately half the members suffer from a severe speech and language disorder inherited as an autosomal dominant trait. Affected members exhibit a striking deficit in orofacial motor control — they struggle to coordinate the rapid, precise movements of the lips, tongue, and palate required for fluent speech — along with broader impairments in grammatical processing and nonverbal cognition.24 Neuroimaging studies revealed that affected KE family members have bilateral reductions in the volume of the caudate nucleus and abnormal patterns of activation in Broca's area and the cerebellum during speech tasks.10
In 2001, Lai and colleagues identified the genetic basis of the KE family's disorder: a point mutation in the FOXP2 gene on chromosome 7, which encodes a transcription factor containing a forkhead DNA-binding domain. The mutation, a guanine-to-adenine transition producing an arginine-to-histidine substitution at position 553 (R553H), disrupts the DNA-binding ability of the protein and cosegregates perfectly with the disorder in the family.7 The following year, Enard and colleagues sequenced FOXP2 across multiple primate species and the mouse, revealing that the human version of the protein differs from that of chimpanzees by two amino acid substitutions, and that the pattern of nucleotide variation around these substitutions is consistent with a recent selective sweep, suggesting that FOXP2 was a target of positive selection in the human lineage within the last 200,000 years.8
In 2007, Krause and colleagues extracted FOXP2 sequences from Neanderthal remains from El Sidrón, Spain, and demonstrated that Neanderthals shared both of the derived amino acid changes found in modern humans.9 This finding implies that the human-derived form of FOXP2 was already present in the last common ancestor of modern humans and Neanderthals, at least 400,000 to 500,000 years ago, pushing back the timeline for this particular genetic change considerably. However, subsequent research has identified regulatory differences between human and Neanderthal FOXP2, and it remains unclear whether the gene functioned identically in both species.9, 22 FOXP2 is now understood not as a "language gene" in isolation but as a transcription factor that regulates the expression of hundreds of downstream genes involved in neural development, synaptic plasticity, and motor learning — making it a critical node in a much larger network.10
Key milestones in FOXP2 research7, 8, 9, 10, 24
| Year | Discovery | Significance |
|---|---|---|
| 1990 | KE family disorder linked to chromosome 7 | First identification of a Mendelian locus for a speech and language disorder |
| 1995 | Detailed phenotyping of KE family | Showed the disorder involves orofacial praxis, grammar, and nonverbal cognition |
| 2001 | FOXP2 gene identified (R553H mutation) | First specific gene implicated in speech and language |
| 2002 | Cross-species FOXP2 comparison | Two human-specific amino acid changes under positive selection |
| 2007 | Neanderthal FOXP2 sequenced | Derived human variant shared with Neanderthals; origin ≥400,000 years ago |
Gestural origins theories
One influential family of hypotheses proposes that language evolved first as a system of manual gestures before transitioning to the vocal-auditory channel. The gestural-origins theory draws on several observations: great apes are far more flexible and intentional in their gestural communication than in their vocalisations, which tend to be emotionally driven and involuntary; the neural circuits for hand control and speech production overlap extensively in the human brain, particularly in Broca's area; and signed languages possess all the structural complexity of spoken languages, demonstrating that the vocal-auditory modality is not inherently necessary for language.12, 25
Michael Corballis developed the gesture-first hypothesis in detail in his 2002 book From Hand to Mouth, arguing that the evolution of bipedalism freed the hands for communicative gesturing and that a manual protolanguage gradually accumulated complexity over millions of years before a relatively late switch to speech, perhaps driven by the advantages of freeing the hands for tool use and allowing communication in the dark or at a distance.12 The neurological foundation for this proposal was strengthened by the discovery of mirror neurons in the macaque premotor cortex by Rizzolatti and colleagues. These neurons fire both when a monkey performs a hand action and when it observes the same action performed by another individual, providing a neural mechanism for understanding the actions and intentions of others.11
Rizzolatti and Arbib proposed in 1998 that the mirror neuron system, located in an area homologous to Broca's area in humans, provided the evolutionary platform from which language emerged, bridging the gap between action recognition and intentional communication.11 Arbib later elaborated this into the mirror system hypothesis, proposing a sequence of evolutionary stages from simple grasping, through complex imitation, to pantomime, protosign, protospeech, and finally full language, with each stage building on neural circuitry established by its predecessor.13 Critics of the gestural theory have noted that the transition from manual to vocal communication remains poorly explained, that vocalisation rather than gesture is the dominant modality of primate communication more broadly, and that the mirror neuron system's role in language remains uncertain in humans.2, 25
Vocal and musical protolanguage theories
An alternative set of hypotheses proposes that language evolved directly through the vocal channel, beginning with a "protolanguage" — a communication system possessing some but not all of the properties of modern language. Vocal protolanguage theories generally envision an early stage in which hominins used individual vocalisations as holistic signals for entire meanings (much as vervet monkeys use distinct alarm calls for different predators), followed by a process of segmentation, combination, and compositional semantics that gradually built up the structured grammar of modern language.2, 25
Steven Mithen proposed a variant of this approach in his 2005 book The Singing Neanderthals, arguing that the ancestral communication system was a holistic, manipulative, multi-modal, musical, and mimetic system he termed "Hmmmmm." In Mithen's account, this musical protolanguage served emotional and social bonding functions — much as music does today — and predated the emergence of compositional, referential language. The separation of music and language from a common ancestor would explain both their deep similarities (both are hierarchically structured sequences of discrete acoustic units) and their differences (music primarily expresses emotion; language primarily conveys propositional content).14
W. Tecumseh Fitch, in his comprehensive 2010 synthesis, evaluated multiple protolanguage models and argued that the evidence best supports a scenario in which a lexical protolanguage — a system of meaningful vocalisations without syntax — preceded the evolution of hierarchical phrase structure. In this view, the crucial innovation was not the capacity for vocalisation per se, which is ancient and widespread among vertebrates, but the evolution of vocal learning — the ability to imitate and modify vocalisations based on auditory input — which in mammals is extremely rare, found only in humans, cetaceans, elephants, pinnipeds, and bats.2 The neural changes underlying vocal learning, including direct cortical control of the laryngeal muscles via the pyramidal tract, may have been among the key adaptations enabling speech in the hominin lineage.2, 25
Social complexity and group size
Robin Dunbar's social brain hypothesis, articulated in a landmark 1993 paper, provides a functional account of why language might have evolved.
{kind=link}
Dunbar observed that across primate species, neocortex size correlates strongly with social group size, and proposed that the primate neocortex expanded primarily to manage the cognitive demands of living in complex social groups — tracking relationships, detecting cheaters, forming alliances, and predicting the behaviour of others.15 Dunbar observed that across primate species, neocortex size correlates strongly with social group size, and proposed that the primate neocortex expanded primarily to manage the cognitive demands of living in complex social groups — tracking relationships, detecting cheaters, forming alliances, and predicting the behaviour of others.15 Extrapolating the primate regression line to the human neocortex ratio predicts a natural group size of approximately 150 individuals, a figure now known as Dunbar's number, which corresponds remarkably well to the size of many real human social units, from Neolithic villages to modern military companies and personal social networks.15
Dunbar further argued that language evolved as a more efficient substitute for social grooming — the physical touching and hair-picking that primates use to maintain social bonds. Grooming is effective but time-consuming: in species with large group sizes, individuals must devote an increasing fraction of their waking hours to grooming partners, and at the group sizes characteristic of humans, the time budget would become unsustainable. Language, Dunbar proposed, allowed early humans to "groom" multiple social partners simultaneously through conversation, and to exchange social information — gossip — about third parties who were not physically present, vastly expanding the range and efficiency of social bonding.15
The social brain hypothesis predicts that the primary selective pressure for language was social rather than technological or ecological, and that the earliest functions of language were social in nature — managing alliances, exchanging gossip, and reinforcing group cohesion — rather than conveying information about tool-making techniques or the locations of food sources. Studies of modern conversational content broadly support this prediction: approximately two-thirds of human conversation is devoted to social topics.15 However, the hypothesis has been criticised for treating "group size" as a monolithic variable and for relying on a correlation across primate species that may not straightforwardly apply to the unique trajectory of human brain evolution.2
The timing debate: gradual emergence versus sudden saltation
One of the most contested questions in language evolution is whether the capacity for language emerged gradually over hundreds of thousands or even millions of years, or appeared relatively suddenly as the result of a discrete genetic or neurological event. The saltation hypothesis is most closely associated with Richard Klein, who argued that the dramatic proliferation of symbolic artefacts, complex technology, and long-distance exchange networks in the European Upper Palaeolithic, beginning approximately 40,000 to 50,000 years ago, reflects a sudden cognitive revolution — possibly triggered by a fortuitous genetic mutation — that brought about fully modern language and with it the full suite of modern human behaviours.16 In Klein's view, anatomically modern humans existed for more than 100,000 years before this revolution, living in ways that were archaeologically indistinguishable from Neanderthals, and the abrupt appearance of symbolic culture marks the moment when language "switched on."16
The gradualist position was forcefully argued by McBrearty and Brooks in their influential 2000 review, which documented that many of the behaviours supposedly diagnostic of Klein's "revolution" — including blade technology, long-distance raw material transport, pigment use, bone tools, and fishing — appear in the African Middle Stone Age tens of thousands of years before their European counterparts, in a piecemeal, cumulative fashion rather than as a sudden package.17 This evidence suggests that modern cognition, and by extension language, assembled gradually in Africa over a period spanning at least 100,000 years, and that the apparent "revolution" in Europe reflects the arrival of populations that already possessed these capacities rather than their sudden invention.17
Dediu and Levinson pushed the gradualist argument further in 2013, arguing on genetic, anatomical, and archaeological grounds that recognisably modern language is likely an ancient feature of the genus Homo, predating at least the last common ancestor of modern humans and Neanderthals approximately 500,000 years ago.22 If Neanderthals shared with modern humans the derived FOXP2 variant, a modern-like hyoid bone, and the capacity for symbolic behaviour (as suggested by their production of ochre, eagle-talon ornaments, and cave markings), then the simplest interpretation is that the common ancestor of both lineages already possessed the biological and cognitive substrate for language.9, 22 Under this view, language has had a long evolutionary history characterised by incremental culture-gene coevolution, rather than a single dramatic mutation or rewiring event.22
Archaeological proxies for symbolic cognition
Because language itself does not preserve in the archaeological record, researchers have sought material proxies for the symbolic thinking that language presumably requires or accompanies. The logic is straightforward: if an artefact can only be explained by the capacity to assign arbitrary meaning to a physical object or pattern — that is, to use symbols — then its maker likely possessed the cognitive architecture that also underlies language.17, 18
{kind=link}
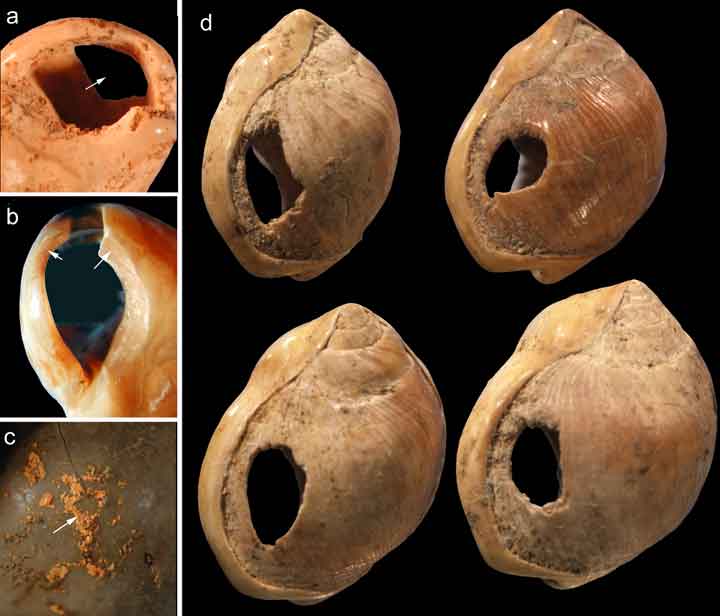
The most important such proxies include the engraved ochre pieces from Blombos Cave in South Africa, dated to approximately 77,000 years ago. These small blocks of red ochre bear deliberately incised cross-hatched patterns that have no obvious utilitarian function and appear to represent intentional geometric designs, making them among the oldest known examples of abstract symbolic expression.18 Perforated marine shell beads, fashioned from Nassarius species, have been recovered from Middle Stone Age and Middle Palaeolithic sites across Africa, including Blombos Cave, and from Grotte des Pigeons at Taforalt in Morocco, where they date to approximately 82,000 years ago.19 The shells show signs of deliberate perforation, use-wear from stringing, and coating with red ochre, indicating that they were worn as personal ornaments — items whose meaning derives from social convention rather than physical function.19
The appearance of figurative art, including the depiction of recognisable animals and human-animal composite figures, is another important proxy. The oldest known figurative cave art includes depictions of pigs and hand stencils from Sulawesi, Indonesia, dated to at least 45,500 years ago, and the Chauvet Cave paintings in France, dated to approximately 36,000 years ago.17 Figurative art implies not only symbolic capacity but also the ability to plan and execute complex representations, to share cultural knowledge about techniques and conventions, and arguably to narrate — to tell stories about the depicted subjects.2, 17 The cumulative picture from the archaeological record is one of a gradual, mosaic accumulation of symbolic behaviours over tens of thousands of years in Africa, broadly consistent with a gradualist model of language evolution and inconsistent with a single, sudden cognitive revolution.17, 18, 19
Universal Grammar and its critics
The theoretical debate over language origins is deeply intertwined with the question of what language is — what its essential structural properties are and how much of its architecture is genetically specified. The most influential answer came from Noam Chomsky, who proposed that all human languages share a common underlying structure, a Universal Grammar (UG), that is innate to the human mind and genetically encoded. In a landmark 2002 paper with Hauser and Fitch, Chomsky distinguished between the faculty of language in the broad sense (FLB), encompassing the sensory-motor and conceptual-intentional systems that are largely shared with other animals, and the faculty of language in the narrow sense (FLN), which they proposed might consist solely of recursion — the capacity to embed phrases within phrases to generate an infinite set of hierarchically structured expressions from a finite set of elements.1
The Hauser-Chomsky-Fitch hypothesis implies that the evolution of language required a single critical innovation — the computational mechanism of recursion — which was then layered onto pre-existing sensory-motor and conceptual systems. If correct, this would make language evolution appear relatively simple from a biological standpoint: a single genetic or developmental change could have produced the recursive capacity, and the rest of language would follow from the integration of this new operation with existing cognitive systems.1
This minimalist view was vigorously challenged by Pinker and Jackendoff in 2005, who argued that many aspects of language beyond recursion are both uniquely human and uniquely linguistic — including phonology (the systematic organisation of speech sounds), morphology (the internal structure of words), the mapping between phonological form and meaning, and the pragmatic principles governing conversation. These components, they argued, require their own evolutionary explanations and cannot be reduced to a single recursive mechanism grafted onto domain-general cognition.20
A different kind of challenge came from usage-based approaches, most influentially articulated by Michael Tomasello. Tomasello argued that children acquire language not by deploying an innate grammar but through domain-general cognitive processes — intention reading, pattern finding, and analogical reasoning — applied to the linguistic input they receive from their community. In this view, the structural regularities of language emerge from the repeated use and transmission of communicative conventions across generations, and the biological endowment for language consists not of a specific grammar module but of the general capacities for shared intentionality, cultural learning, and cooperative communication that distinguish humans from other primates.21 The debate between nativist and usage-based accounts remains one of the central fault lines in linguistic theory, with direct implications for how the evolution of language is understood.1, 20, 21
Language, culture, and cumulative cultural evolution
Whatever its precise origins, language is inseparable from the broader phenomenon of cumulative cultural evolution — the process by which cultural knowledge is transmitted, modified, and accumulated across generations, producing technologies, institutions, and bodies of knowledge that no single individual could invent independently. Language is both a product and an engine of this process: it is itself a culturally transmitted system that changes over generations, and it is also the primary medium through which all other cultural knowledge is transmitted, from tool-making techniques to social norms to cosmological beliefs.2, 23
Henrich demonstrated formally in 2004 that population size and interconnectedness are critical variables for cumulative cultural evolution: larger, better-connected populations can sustain more complex cultural repertoires because they provide a larger pool of innovators and a more reliable system of social transmission, reducing the probability that useful knowledge is lost through random demographic fluctuations.23 Language would have amplified this process enormously, allowing precise information to be transmitted with high fidelity across individuals and generations, enabling the teaching of complex skills, and facilitating the coordination of large-scale cooperative enterprises such as communal hunting, inter-group trade, and territorial defence.2, 23
This suggests a feedback loop in which language and culture coevolved: the emergence of even a rudimentary protolanguage would have enhanced the fidelity of cultural transmission, which in turn would have increased the adaptive value of more complex language, creating selective pressure for the biological capacities that support it — finer vocal control, larger working memory, more sophisticated social cognition.2, 22 The result is a system in which biological evolution and cultural evolution are intertwined in a spiral of mutual reinforcement, each driving the elaboration of the other. This culture-gene coevolutionary perspective, increasingly supported by evidence from genetics, archaeology, and comparative psychology, may offer the most promising framework for understanding how the extraordinary human capacity for language could have emerged from the incremental, undirected process of natural selection.22, 23, 25
References
The revolution that wasn't: a new interpretation of the origin of modern human behavior
82,000-year-old shell beads from North Africa and implications for the origins of modern human behavior
On the antiquity of language: the reinterpretation of Neandertal linguistic capacities and its consequences
Demography and cultural evolution: how adaptive cultural processes produced maladaptive losses in Tasmania
Praxic and nonverbal cognitive deficits in a large family with a genetically transmitted speech and language disorder