Overview
- Multiple waves of hominin dispersal have left Africa over the past two million years, beginning with Homo erectus at Dmanisi in Georgia by 1.85 million years ago and culminating in the main Homo sapiens expansion roughly 60,000–70,000 years ago that seeded every inhabited continent.
- Genetic evidence from mitochondrial DNA, Y-chromosome lineages, and whole-genome analyses reveals a serial founder effect model in which human genetic diversity declines systematically with geographic distance from Africa, consistent with successive small founding populations branching off during the global dispersal.
- As modern humans spread across Eurasia and Oceania, they interbred with at least two archaic hominin populations — Neanderthals and Denisovans — leaving signatures of admixture that persist in non-African genomes today and that have contributed functionally important genetic variants to living human populations.
The dispersal of human populations out of Africa is one of the defining narratives in the history of life on Earth. Over the past two million years, at least three major hominin lineages have expanded beyond the African continent, each under different ecological conditions and at different stages of biological and cultural evolution. The earliest of these dispersals carried Homo erectus into the Caucasus and across southern Asia by approximately 1.85 million years ago.1 A later expansion brought Homo heidelbergensis and related populations into Europe and western Asia during the Middle Pleistocene.3 The most consequential dispersal, however, was the spread of anatomically modern Homo sapiens out of Africa roughly 60,000 to 70,000 years ago, an event that ultimately led to human settlement of every habitable landmass on the planet, from the tropical coasts of Southeast Asia and Australia to the ice-age steppes of northern Europe and the Americas.4, 5, 6
The evidence for these dispersals comes from multiple, independent lines of inquiry: the fossil and archaeological record, mitochondrial and Y-chromosome phylogeography, whole-genome sequencing of both living and ancient populations, and palaeoclimatic reconstructions of the corridors through which migration was possible. Together, these data reveal a complex history of expansion, admixture with archaic populations encountered along the way, and adaptation to radically new environments, shaped at every stage by the oscillations of Pleistocene climate.8, 12, 22
Early hominin dispersals
The earliest well-dated evidence of hominins outside Africa comes from the site of Dmanisi in the Republic of Georgia. Archaeological excavations and palaeomagnetic dating have established that hominins occupied Dmanisi between approximately 1.85 and 1.78 million years ago, making it the oldest securely dated hominin site in Eurasia.1 The Dmanisi fossils represent small-bodied, small-brained individuals assigned to early Homo erectus (sometimes designated Homo erectus georgicus), with cranial capacities ranging from roughly 546 to 730 cubic centimetres. A remarkably complete skull described in 2013, known as Skull 5, displayed a combination of a small braincase and a large, projecting face that challenged assumptions about the degree of morphological variation within early Homo, leading some researchers to argue that several species previously recognised in the early Pleistocene fossil record of Africa may in fact represent a single, highly variable lineage.2

{kind=link}
The Dmanisi hominins carried simple Oldowan-type stone tools rather than the more sophisticated Acheulean handaxes associated with later H. erectus populations in Africa, indicating that the initial expansion out of Africa did not require advanced lithic technology.1, 2 The occupation of Dmanisi appears to have been facilitated by the site's location at the crossroads of Africa and Asia, in a temperate grassland environment that supported abundant large mammals. The Dmanisi evidence demonstrates that the first hominin dispersal out of Africa occurred at the same time as, or even slightly before, the earliest appearance of classic H. erectus in the East African fossil record, raising the possibility that the genus Homo diversified across a broad geographic range rather than in a single African centre of origin.1
Following the Dmanisi expansion, Homo erectus populations spread widely across southern and eastern Asia. Fossils attributed to H. erectus in Java (Indonesia) have been dated to approximately 1.5 to 1.0 million years ago, and the species persisted in island Southeast Asia until as recently as 100,000 years ago in some estimates.3 A subsequent major dispersal occurred during the Middle Pleistocene, when populations of Homo heidelbergensis (or closely related archaic humans) expanded from Africa into Europe and western Asia by approximately 600,000 to 700,000 years ago, carrying Acheulean stone tool technology and establishing the lineage that would eventually give rise to the Neanderthals in Europe and the Denisovans in Asia.3
The main Homo sapiens dispersal
.png)
.png){kind=link}
The dispersal that reshaped the biological and cultural landscape of the entire planet was the expansion of anatomically modern Homo sapiens out of Africa. Genetic evidence places this event at approximately 60,000 to 70,000 years ago. The mitochondrial DNA phylogeny is particularly informative: all non-African mitochondrial lineages descend from haplogroup L3, which arose in East Africa at roughly 70,000 years ago. The near-identity in age between L3 and its two non-African daughter haplogroups, M and N, suggests that the expansion within eastern Africa and the dispersal out of the continent were part of the same demographic process, with a small founding population carrying L3 derivatives through the gateway to Asia.5
The precise route of this dispersal remains debated. The southern coastal route hypothesis proposes that modern humans crossed the southern end of the Red Sea at the Bab el-Mandeb strait during a period of low sea level and then moved rapidly eastward along the coasts of the Arabian Peninsula, the Indian subcontinent, and Southeast Asia, exploiting marine and littoral resources as they went.6 Archaeological evidence from Jebel Faya in the United Arab Emirates, where stone tools with affinities to the African Middle Stone Age have been dated to approximately 125,000 to 100,000 years ago, suggests that anatomically modern humans may have been present in Arabia significantly earlier than the main 60,000-year-ago expansion, possibly during an earlier, ultimately less successful dispersal event during the last interglacial.7
An alternative or complementary model emphasises the northern Levantine corridor, through the Sinai Peninsula and into the Levant, as the primary exit route. The two models are not mutually exclusive, and growing evidence suggests that multiple dispersals occurred at different times and by different paths, with only a subset leaving a lasting genetic imprint on living non-African populations.8 Palaeoclimatic reconstructions have identified periodic "green corridors" across the Sahara and Arabian Peninsula during humid phases driven by orbital precession cycles, when increased monsoon rainfall transformed desert barriers into habitable savanna and grassland. These humid windows, recurring roughly every 21,000 years, opened and closed migration corridors that would have controlled the timing and feasibility of human passage through otherwise impassable terrain.22
Colonization of Australia and Europe
The southern coastal dispersal carried modern humans to the far reaches of maritime Southeast Asia and, ultimately, to the continent of Sahul (the Pleistocene landmass comprising Australia, New Guinea, and Tasmania, connected when sea levels were lower). Optical stimulated luminescence dating of sediments at the rock shelter of Madjedbebe in northern Australia has yielded ages of approximately 65,000 years for the earliest occupation horizon, which contained a rich assemblage of stone tools including ground-edge hatchet heads, grinding stones, and processed ochre pigments.9 If these dates are correct, the colonization of Australia occurred remarkably soon after the main out-of-Africa event, implying rapid movement along the southern Asian coastline and the capacity for repeated sea crossings, since even at glacial-maximum sea-level lows, the passage from Sunda (mainland Southeast Asia) to Sahul required multiple open-water voyages of at least 90 kilometres.6, 9
The colonization of Europe followed a different trajectory. The earliest evidence of Homo sapiens in Europe comes from Bacho Kiro Cave in Bulgaria, where human remains directly dated to approximately 45,000 years ago were found in association with Initial Upper Palaeolithic (IUP) stone tools, including pointed blades manufactured from high-quality flint sourced up to 180 kilometres from the site.10 A partial skull from Zlatý kůň in the Czech Republic and remains from Ilsenhöhle in Ranis, Germany, have extended the evidence for early H. sapiens presence in the higher latitudes of Europe to around 45,000 years ago as well, demonstrating that modern humans reached central and northern Europe within a few thousand years of their first appearance in the Balkans.24 These pioneer populations coexisted with Neanderthals for several thousand years before the latter disappeared from the European record by approximately 40,000 years ago. The IUP technology that accompanied the earliest European modern humans appears to have originated in southwest Asia and spread rapidly across the mid-latitudes of Eurasia, from the Balkans to Mongolia, reflecting a swift and far-ranging dispersal.10
Peopling of the Americas
The colonization of the Americas was the last major episode in the global dispersal of Homo sapiens, and its timing remains one of the most actively debated questions in archaeology and genetics. During glacial periods of the Late Pleistocene, lowered sea levels exposed the Bering Land Bridge (Beringia), a broad landmass connecting northeastern Siberia to Alaska that was up to 1,600 kilometres wide at its maximum extent. Human populations from northeastern Asia entered eastern Beringia, but the routes by which they penetrated southward into the Americas are contested.17
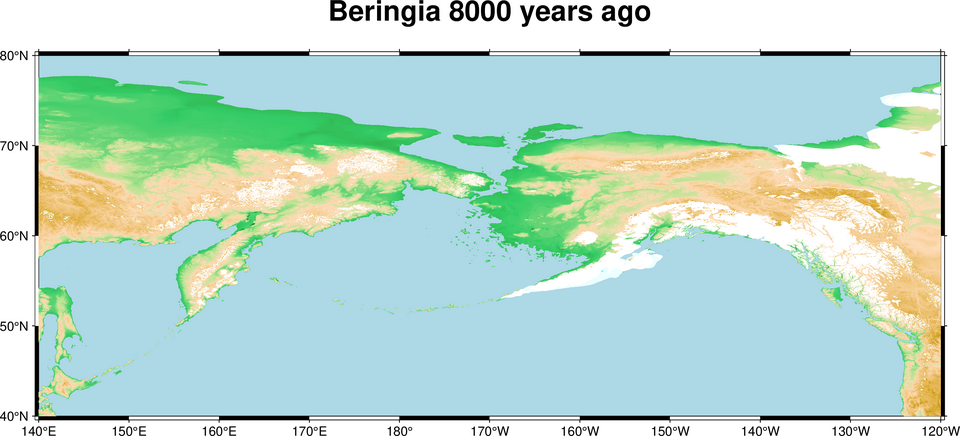
{kind=link}
For decades, the Clovis-first model held that the initial colonizers entered the interior of North America through an ice-free corridor between the Laurentide and Cordilleran ice sheets approximately 13,000 years ago, producing the distinctive fluted Clovis projectile points found across much of the continent. However, environmental DNA and pollen analyses of lake sediment cores from a bottleneck section of the corridor have demonstrated that it did not become biologically viable — with sufficient vegetation, bison, and other game to support human passage — until approximately 12,600 calendar years before present, too late to account for pre-Clovis sites already documented farther south.17
The accumulating evidence for pre-Clovis occupation has shifted scholarly consensus toward a Pacific coastal route, in which populations moved southward by boat along the coast of Beringia and down the Pacific margin, exploiting kelp-forest ecosystems and littoral resources. The site of Monte Verde in southern Chile, excavated by Tom Dillehay and colleagues, provides some of the most robust pre-Clovis evidence: multiple occupation horizons with directly dated artefacts and organic remains extend back to at least 14,500 years ago, with additional finds suggesting human presence as early as 18,500 years ago.18 Human footprints discovered at White Sands, New Mexico, have been dated by multiple independent methods — radiocarbon dating of plant seeds, pollen, and ancient lakebed sediment — to between approximately 23,000 and 21,000 years ago, placing humans in interior North America during the Last Glacial Maximum, far earlier than any version of the Clovis-first model allows.16
Genomic analyses of ancient and living Native American populations have further refined the picture. Ancient DNA from individuals spanning Alaska to Patagonia reveals an early split within Beringian populations into Northern and Southern lineages, followed by rapid southward dispersal and diversification that included multiple independent migration events.19 The genetic data are consistent with an initial entry into the Americas before approximately 16,000 years ago, with subsequent population movements producing the complex mosaic of Native American genetic diversity observed today.19
Major dispersal events in human global expansion1, 5, 9, 10, 16, 20
| Dispersal event | Approximate date | Route or destination | Key evidence |
|---|---|---|---|
| Homo erectus out of Africa | ~1.85 Ma | Levant → Caucasus → southern Asia | Dmanisi fossils and Oldowan tools |
| H. heidelbergensis dispersal | ~700–600 ka | Africa → Europe & western Asia | Acheulean tools; Mauer, Bodo fossils |
| H. sapiens main out-of-Africa | ~70–60 ka | Southern coastal &/or Levantine route | mtDNA haplogroup L3; M and N lineages |
| Colonization of Australia (Sahul) | ~65 ka | Sunda → Sahul via sea crossings | Madjedbebe rock shelter artefacts |
| Colonization of Europe | ~45 ka | Southwest Asia → Balkans → central Europe | Bacho Kiro Cave; Ranis; Zlatý kůň |
| Peopling of the Americas | ~23–16 ka | Beringia → Pacific coastal route | White Sands footprints; Monte Verde |
| Remote Oceania (Polynesian expansion) | ~3,000–800 ya | Island Southeast Asia → Pacific islands | Lapita pottery; ancient DNA from Vanuatu and Tonga |
The Polynesian expansion
The settlement of Remote Oceania — the vast Pacific island region east of the Solomon Islands, including Vanuatu, Fiji, Tonga, Samoa, and ultimately the far-flung islands of Polynesia — represents the last great chapter in the human colonization of the Earth. This expansion was accomplished by Austronesian-speaking peoples whose ultimate origins trace to Taiwan and the coast of southern China, where the Austronesian linguistic family began to diversify approximately 5,000 to 4,000 years ago. Bearers of the Lapita cultural complex, characterised by distinctive dentate-stamped pottery, appeared in the Bismarck Archipelago by approximately 3,300 years ago and rapidly colonized the islands of Vanuatu, New Caledonia, Fiji, Tonga, and Samoa over the following centuries.20
Ancient DNA from early Lapita-associated burials in Vanuatu and Tonga, dating to approximately 3,100 to 2,300 years ago, reveals that the first Remote Oceanians derived almost entirely from East Asian ancestry, with little or no genetic contribution from the Papuan-speaking populations of Near Oceania who had occupied New Guinea and the Solomon Islands for tens of thousands of years prior.20 The substantial Papuan ancestry observed in modern Polynesian populations must therefore reflect later admixture events, in which Papuan-related groups expanded into islands already settled by Lapita descendants and the two populations mixed over subsequent millennia.
From their initial staging area in western Polynesia, Austronesian voyagers embarked on the colonization of the central and eastern Pacific: the Marquesas by approximately 1,000 years ago, Hawai'i and Rapa Nui (Easter Island) by approximately 800 years ago, and New Zealand — the last major landmass on Earth to be permanently settled by humans — by approximately 700 to 750 years ago.20 These voyages, undertaken in double-hulled canoes navigated by stellar, wave, and current observations across thousands of kilometres of open ocean, rank among the most remarkable feats of exploration in human history.
Genetic evidence for dispersal
The reconstruction of human dispersal patterns has been transformed by molecular genetics, beginning with the landmark 1987 study by Rebecca Cann, Mark Stoneking, and Allan Wilson, which analysed mitochondrial DNA from 147 individuals drawn from five geographic populations. The resulting phylogenetic tree placed the deepest-rooting branches in Africa and estimated a coalescence time of approximately 200,000 years for all human mitochondrial lineages, providing the first genetic support for the hypothesis that all living humans descend from a recent African ancestor — a concept subsequently known in popular discourse as "Mitochondrial Eve."4
The mitochondrial phylogeny has since been refined with complete mitogenome sequences from thousands of individuals. The deepest branches of the tree, haplogroups L0 through L6, are found exclusively or predominantly in Africa, while all non-African lineages fall within haplogroups M and N, both of which descend from haplogroup L3.5 The Y-chromosome phylogeny tells a parallel story: the most basal Y-chromosome lineages (haplogroups A and B) are confined to Africa, and all non-African Y-chromosome diversity descends from haplogroups derived from the African tree, consistent with a single primary origin on the continent.23
Whole-genome data from globally distributed populations have revealed a striking geographic pattern: heterozygosity (a measure of genetic diversity within a population) is highest in sub-Saharan Africa and declines progressively and approximately linearly with geographic distance from the continent, a pattern predicted by a serial founder effect model. In this model, each successive colonizing population was founded by a small subset of the population at the expanding front, carrying only a fraction of the genetic diversity of its parent population. The result is a stepwise reduction in diversity with each founding event, producing the observed global gradient.11 The 1000 Genomes Project, which catalogued genetic variation from 2,504 individuals across 26 populations, confirmed this pattern at whole-genome resolution and provided a detailed atlas of population structure, admixture, and demographic history for populations worldwide.21
Decline in genetic diversity with distance from Africa (serial founder effect)11, 23
Admixture with archaic humans
One of the most transformative discoveries of the genomic era is that the global dispersal of Homo sapiens was accompanied by interbreeding with at least two archaic hominin populations. The sequencing of the Neanderthal genome in 2010 by the team led by Svante Paabo revealed that all non-African modern humans carry approximately 1 to 4 percent Neanderthal-derived DNA, indicating that hybridisation occurred when the expanding modern human population encountered Neanderthals, most likely in the Middle East shortly after the exit from Africa.12 Subsequent analyses have mapped the "landscape" of Neanderthal ancestry across the modern human genome, finding that introgressed Neanderthal alleles are enriched in genomic regions influencing keratin filament biology — proteins important for skin and hair, suggesting that Neanderthal genes helped modern humans adapt to colder, non-African environments — while being depleted on the X chromosome and in genes highly expressed in the testes, indicative of reduced fertility in male hybrids.14
In the same year, the genome of a previously unknown archaic population, the Denisovans, was recovered from a finger bone discovered in Denisova Cave in the Altai Mountains of Siberia.13 Denisovan DNA is absent or present at only trace levels in most modern human populations, but it constitutes approximately 4 to 6 percent of the genomes of Melanesian and Aboriginal Australian populations, indicating that modern humans interbred with Denisovans as they moved through Southeast Asia on their way to Oceania.13 Remarkably, a 2021 study of 118 ethnic groups in the Philippines found that the Ayta Magbukon people possess the highest level of Denisovan ancestry in the world — approximately 30 to 40 percent more than Australo-Papuan populations — consistent with an independent admixture event on the Philippine islands followed by relative genetic isolation that preserved the archaic signal.15
The discovery that multiple episodes of archaic admixture accompanied the out-of-Africa dispersal has fundamentally altered the picture of recent human evolution. Rather than a simple replacement of archaic populations by incoming modern humans, the process was one of assimilation with admixture: modern humans absorbed small but genetically consequential contributions from the archaic populations they encountered, and some of these introgressed alleles have been positively selected because they conferred adaptive advantages in new environments. Denisovan-derived variants in the gene EPAS1, for example, are found at high frequency in Tibetan populations and contribute to their ability to tolerate hypoxia at high altitude — a trait inherited not from their own recent ancestors but from an archaic lineage that had already adapted to the conditions of the Tibetan Plateau or its surroundings.14
Climate as a driver of migration corridors
The timing and routes of human dispersals out of Africa were profoundly shaped by Pleistocene climate variability. The Sahara Desert, which today constitutes a formidable barrier between sub-Saharan Africa and the Mediterranean world, has alternated repeatedly between hyperarid desert and habitable "green Sahara" conditions in response to variations in Earth's orbital precession. Approximately every 21,000 years, precessional forcing intensifies the West African monsoon, shifting the tropical rain belt northward and transforming the Sahara into a mosaic of lakes, rivers, and grassland that supports large mammal faunas and, critically, opens corridors for human movement between tropical Africa and North Africa.22
Geochemical analysis of marine sediment cores from the eastern Mediterranean has identified a humid corridor stretching across the Sahara approximately 120,000 years ago, during Marine Isotope Stage 5, which coincides with archaeological evidence of modern human presence in the Levant at sites such as the caves of Skhul and Qafzeh in present-day Israel.22 This early foray into the Levant appears not to have resulted in a permanent colonization of Eurasia, as the archaeological and genetic record suggests that these populations either went extinct or retreated back into Africa as the Saharan corridor closed during subsequent arid phases. The successful, lasting expansion at 60,000 to 70,000 years ago may have coincided with a later humid interval that once again opened the passage, although the precise environmental conditions remain a subject of active investigation.8
Climate also shaped later dispersals. The colonization of northern Europe required adaptation to intensely cold, seasonal environments during Marine Isotope Stage 3, when the presence of Neanderthals already demonstrated that hominin occupation of these latitudes was possible but ecologically demanding.10, 24 The peopling of the Americas was gated by the extent of the Beringian land bridge, which was exposed during glacial periods of lowered sea level, and by the opening of viable southward routes, whether along the Pacific coast or through the interior ice-free corridor, that were themselves controlled by the advance and retreat of continental ice sheets.17 In the Pacific, the Polynesian expansion appears to have been facilitated in part by the stable climatic conditions and predictable trade-wind patterns of the late Holocene, which made long-distance oceanic voyaging practical with the watercraft technology available to Austronesian peoples.20
The human story, taken as a whole, is one of repeated interaction between biological capacity and environmental opportunity. Each dispersal required a population with sufficient demographic momentum, cultural technology, and ecological flexibility to exploit corridors that opened and closed on timescales of thousands to tens of thousands of years. The genetic signatures of these passages — founder effects, admixture gradients, and adaptive introgression from archaic populations — remain written in the genomes of living people, providing an increasingly detailed record of how a single African species came to inhabit every corner of the globe.11, 21, 23
References
Going east: new genetic and archaeological perspectives on the modern human colonization of Eurasia
The southern route ‘out of Africa’: evidence for an early expansion of modern humans into Arabia
Support from the relationship of genetic and geographic distance in human populations for a serial founder effect originating in Africa
Evidence for human footprints in White Sands, New Mexico dating to the Last Glacial Maximum
A humid corridor across the Sahara for the migration of early modern humans out of Africa 120,000 years ago