Overview
- The claim that there are no transitional fossils between apes and humans is contradicted by a detailed morphological sequence spanning more than 7 million years, from Sahelanthropus through Ardipithecus, Australopithecus, early Homo, H. erectus, and H. heidelbergensis to anatomically modern humans.
- Creationist attempts to sort fossil hominins into discrete ‘ape’ or ‘human’ categories consistently fail because the gradations in brain size, facial anatomy, dentition, and locomotion are continuous—different creationist organizations cannot even agree on which side of the line to place the same specimens.
- Genetic evidence independently corroborates the fossil record: the fusion scar on human chromosome 2, shared endogenous retroviruses in identical genomic positions across great apes, and the shared broken GULO pseudogene all point to common ancestry and make independent origins of humans effectively impossible.
Few topics in science have been more systematically misrepresented in popular discourse than the human fossil record. Since the publication of Charles Darwin’s The Descent of Man in 1871—in which Darwin predicted, on purely anatomical and biogeographical grounds, that humanity’s origins would prove to lie in Africa10—paleoanthropologists have recovered hundreds of hominin specimens documenting a continuous morphological progression across more than seven million years. Yet creationist organizations continue to assert that this record is absent, ambiguous, or fraudulent. Each of these claims can be examined against the evidence.
{kind=link}
This article addresses the principal creationist objections to human evolution in the order they most commonly appear: the claim that transitional fossils are absent, the categorical argument that each specimen must be either a full ape or a full human, specific attacks on individual fossils, and the independent genetic evidence that confirms common ancestry. The goal is not polemic but accurate summary of what the fossil and genomic records actually show.
The transitional fossil claim
The most frequently repeated creationist claim about human origins is that there are no transitional fossils bridging apes and humans. In its strongest form—associated with writers such as Casey Luskin of the Discovery Institute and the editorial team at Answers in Genesis—the assertion is that the gap between non-human apes and anatomically modern humans is unbridged by any intermediate forms.12, 13 The hominin fossil record directly contradicts this.
Beginning at the base of the human lineage, Sahelanthropus tchadensis, dated to approximately 7 million years ago from the Djurab Desert of Chad, combines a chimpanzee-sized braincase of roughly 360 cubic centimeters with small, non-honing canine teeth and a foramen magnum positioned in a manner consistent with upright posture—features that place it close to or on the hominin lineage.1 Roughly a million years later, Orrorin tugenensis from Kenya preserves femoral morphology interpreted as evidence of bipedalism, though the debate over its locomotion continues.12
By 4.4 million years ago, Ardipithecus ramidus is documented by a nearly complete skeleton from Ethiopia. Ardipithecus retains a divergent, grasping big toe suited for arboreal movement yet also shows pelvic and lower-limb anatomy indicating facultative bipedalism on the ground—a mosaic of features that makes it a genuine intermediate between arboreal apes and the obligate bipeds that would follow.2 Its canines are reduced compared to those of chimpanzees and gorillas, its face is less projecting, and its brain, at roughly 300–350 cubic centimeters, is ape-sized. No paleoanthropologist classifies Ardipithecus as either a modern ape or a modern human; it occupies exactly the kind of intermediate position that the transitional fossil claim says cannot exist.
The australopithecines that follow—Australopithecus anamensis, A. afarensis, A. africanus, and related species from 4.2 to 2.0 million years ago—are fully committed bipeds with small brains (370–550 cc), large back teeth, and projecting faces. They are neither apes nor humans in the colloquial sense; they walked upright like humans but had ape-sized brains and ape-like craniofacial anatomy.3, 15 The famous “Lucy” skeleton (A. afarensis, AL 288-1) preserves the angled femur, valgus knee, and shortened iliac blade of an obligate biped despite a braincase little larger than that of a chimpanzee.3
Early Homo, appearing by 2.8 million years ago at Ledi-Geraru, Ethiopia, shows a chin region and tooth-row reduction more derived than any australopithecine.4 Homo habilis and H. rudolfensis span brain sizes of roughly 510–750 cc and are associated with the earliest stone tools. Homo erectus, emerging by 1.9 million years ago, achieves modern body proportions, a substantially larger brain (750–1,250 cc), and disperses out of Africa into Eurasia.5 Homo heidelbergensis, broadly defined, occupies the interval 700,000–200,000 years ago with brain sizes overlapping the modern human range (1,100–1,400 cc), yet retains prominent brow ridges and other archaic features that distinguish it from anatomically modern H. sapiens.6
The progression from Sahelanthropus through Ardipithecus, Australopithecus, early Homo, H. erectus, H. heidelbergensis, to anatomically modern humans represents a detailed morphological sequence spanning every major character that distinguishes humans from other great apes: upright posture, reduced canines, expanded braincase, reduced brow ridgeing, and modern limb proportions. Each transition is documented by multiple specimens across multiple sites and continents. The hominin fossil record is, by any reasonable measure, among the best-documented vertebrate lineages in the entire fossil record.12, 13
The “ape or human” dichotomy
A second creationist strategy, prominent in the work of Luskin and in publications from both Answers in Genesis and the Discovery Institute, is to argue that each fossil hominin must be assigned to one of only two categories: a non-human ape or a fully modern human. Because science recognizes a continuous gradation, this strategy inevitably produces contradictions. The most telling symptom is that creationist organizations cannot agree on which specimens belong in which category.12, 13
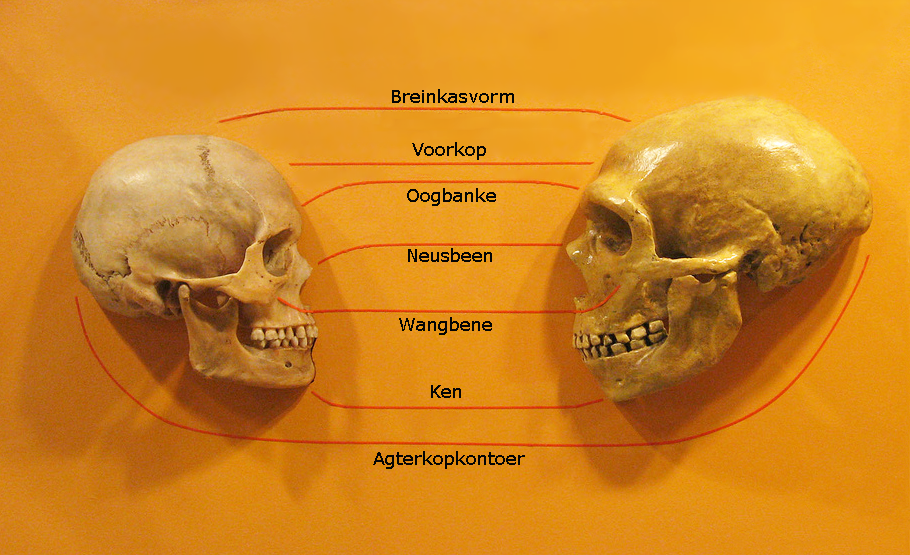
Australopithecus is frequently dismissed as “just an ape” by Answers in Genesis, which argues that australopithecines were knuckle-walkers akin to modern chimpanzees. Yet the postcranial evidence—the angled femur, arched foot, broad pelvis, and foramen magnum position of specimens like Lucy—contradicts knuckle-walking anatomy directly and unambiguously.3, 15 Independent analyses of australopithecine footprints at Laetoli confirm a fully upright gait with no midtarsal break, a feature absent in any living ape.15
Homo habilis creates a different problem for the same framework. Some creationist writers accept it as human because the genus name is Homo; others reject it as an ape because its brain is small and its face is projecting. The Discovery Institute has at various times assigned H. habilis to both categories in different publications. Homo erectus is generally placed in the “human” column by creationists, yet its brow ridges, low cranial vault, and pronounced prognathism differ from modern humans as much as they differ from australopithecines in other directions. The assignment is motivated by category membership rather than morphology.5, 12
The reason creationist taxonomies are internally inconsistent is that the morphological characters themselves change continuously across the fossil sequence.13 Brain volume in hominins increases from roughly 300 cc in Ardipithecus to over 1,400 cc in some H. sapiens specimens, with every intermediate value documented by real fossils. Face projection, brow ridge size, tooth dimensions, and postcranial proportions all change incrementally. There is no principled anatomical threshold at which an ape becomes a human; the dichotomy is imposed on the evidence rather than derived from it.
Specific fossil claims
Creationist literature repeatedly invokes three specific cases as evidence that the human fossil record cannot be trusted: Nebraska Man, Piltdown Man, and the claim that Lucy was “just a chimp.” Each deserves a direct response.11, 17
Nebraska Man (Hesperopithecus haroldcookii) is a favorite example of alleged scientific credulity. In 1922, Henry Fairfield Osborn of the American Museum of Natural History tentatively identified a single worn molar from Nebraska as belonging to an early hominin. Subsequent excavation of the same site in 1927 recovered additional teeth and skeletal material that unambiguously identified the tooth as belonging to a peccary, a pig-like animal. The correction was published promptly by the scientists themselves.17 The episode is cited by creationists as proof that paleoanthropologists reconstruct entire “missing links” from a single tooth, but the historical record shows the opposite: the scientific community identified the error through continued fieldwork and corrected it in the literature within five years. The Nebraska Man episode illustrates how science self-corrects, not how it fails.17
Piltdown Man (Eoanthropus dawsoni), assembled from a human skull cap and an orangutan jaw by Charles Dawson around 1912, is the most notorious fraud in paleoanthropology’s history. Creationists regularly present it as evidence of systematic scientific dishonesty or of willingness to accept fabricated evidence for evolution. The historical record again contradicts this narrative. Many anatomists were skeptical of Piltdown from the outset, noting the mismatch between the skull and jaw. By 1949–1953, fluorine dating and physical examination by a team including Joseph Weiner, Wilfrid Le Gros Clark, and Kenneth Oakley definitively exposed the fraud as a deliberate hoax, almost certainly perpetrated by Dawson himself.11 Piltdown was not exposed by creationists; it was exposed by evolutionary scientists applying rigorous methodology. Its exposure strengthened, not weakened, the scientific case for human evolution, because the genuine hominin fossils—Australopithecus from South Africa, Homo erectus from Java and China—had by then accumulated to the point where Piltdown’s strange anatomy was clearly anomalous.11, 12
Lucy, the 3.2-million-year-old Australopithecus afarensis skeleton (AL 288-1), is periodically claimed by creationist sources to have been “just a chimp” or a tree-dwelling ape with no relevance to human ancestry. This claim focuses on Lucy’s small brain and some features of the wrist and shoulder that have been interpreted as consistent with occasional arboreal activity. It ignores the postcranial evidence that directly addresses terrestrial locomotion: the valgus angle of the femur (the inward angling of the thigh bone that directs weight over the foot), the shortened and broadened ilium of the pelvis, the non-opposable great toe as confirmed by the Laetoli trackways, and the proportions of the knee joint.3, 15 No chimpanzee or gorilla possesses any of these features. Lucy is not a modern human, but she is unambiguously a biped, and no anatomy paper has ever classified A. afarensis postcranials as consistent with knuckle-walking locomotion.3
Darwin’s Africa prediction
One measure of a scientific theory’s power is its ability to generate predictions that are subsequently confirmed by independent evidence. The theory of human evolution by common descent has done this repeatedly, and one of its most striking confirmations involves Africa.10
In The Descent of Man (1871), Darwin wrote that because humans’ closest living relatives—chimpanzees and gorillas—are confined to Africa, it was probable that our earliest ancestors had also lived there.10 At the time Darwin wrote these words, not a single hominin fossil had been found anywhere in Africa. The prediction was based entirely on the anatomical similarity between humans and African apes. It was ridiculed by contemporaries who expected human origins in Europe or Asia.
Between 1924 and the present, every early hominin species—Sahelanthropus, Orrorin, Ardipithecus, Australopithecus, Paranthropus, and the earliest members of Homo—has been found in Africa, precisely as Darwin predicted.1, 2, 4, 12 The oldest hominin fossils outside Africa (H. erectus at Dmanisi, Georgia, dated to 1.77 million years ago) postdate the African H. erectus record, consistent with an African origin followed by dispersal.16 This is the expected pattern under the evolutionary hypothesis, not under the hypothesis of independent creation. The Africa prediction is one of the clearest examples in all of biology of a theoretical prediction confirmed by subsequent empirical discovery.
Completeness of the record
A persistent framing in creationist literature is that the hominin fossil record is “surprisingly sparse” or “fragmentary,” consisting of only a handful of contested specimens. This misrepresents the actual scale of the database. As of the mid-2020s, the hominin fossil record comprises thousands of individual specimens from over a hundred sites across Africa, Europe, and Asia, representing more than twenty recognized species or subspecies.12, 13 Individual species are known from multiple populations across wide geographic ranges: Homo erectus specimens span from East Africa to Indonesia and China, documented across nearly two million years.5, 16
The apparent sparseness of the early hominin record relative to, for example, marine invertebrates reflects the taphonomic reality that large terrestrial vertebrates living in specific forest-edge and riparian environments have low fossilization potential. Bone preservation requires rapid burial under anoxic conditions, and the East African rift system—where most early hominins are found—provides intermittent lacustrine and fluvial environments favorable for preservation only in certain localities.12 Given these constraints, the richness of the hominin fossil record is remarkable, not deficient. Paleoanthropologists frequently note that the major morphological transitions are documented by fossils, and new discoveries consistently fill rather than widen the gaps.
The progressive nature of the discoveries is itself significant. Fossil finds made after theoretical predictions have been formulated provide a stronger test than fossils known before the theory was developed. The discovery of Ardipithecus confirmed predictions about the character states expected in a hominin more primitive than Australopithecus: reduced canines, some degree of bipedalism, and a brain not yet expanded beyond ape range.2 The discovery of early Homo at 2.8 million years ago at Ledi-Geraru filled a temporal gap between late australopithecines and the better-known early Homo assemblages from Olduvai Gorge, again consistent with prior expectation.4
Genetic evidence
Entirely independent of the fossil record, three categories of genomic evidence converge on the same conclusion: humans share common ancestry with other great apes and did not arise by separate creation. These are the chromosome 2 fusion, shared endogenous retroviruses, and the broken GULO pseudogene.7, 8, 9
Human beings have 46 chromosomes (23 pairs), while chimpanzees, gorillas, and orangutans all have 48 (24 pairs). If humans and chimpanzees share a common ancestor, evolutionary theory predicts that at some point in the human lineage, two ancestral chromosomes must have fused end-to-end. Human chromosome 2 carries exactly this signature: it contains two vestigial telomere sequences (which normally occur only at chromosome ends) fused together at its midpoint, along with a remnant centromere corresponding to one of the two ancestral chromosomes, now inactivated.7 The fusion site, characterized in detail by Ijdo and colleagues in 1991, contains the degenerate sequence expected of a head-to-head telomeric fusion.7 No mechanism other than chromosomal fusion produces this configuration, and the configuration aligns precisely with what the common ancestry hypothesis requires.
Endogenous retroviruses (ERVs) are remnants of ancient retroviral infections integrated into the germline DNA of vertebrates. Because retroviral integration into a specific genomic location is an essentially random event, finding the same ERV inserted at the same chromosomal position in two different species is strong evidence that the insertion occurred in a shared ancestor and was inherited by both lineages. The human genome contains approximately 8% ERV-derived sequence.8 Humans and chimpanzees share hundreds of ERVs at identical genomic positions; humans and gorillas share a subset of these; and humans, chimpanzees, gorillas, and orangutans share a further subset.14 This nested pattern of shared ERV insertions precisely mirrors the phylogenetic tree inferred from morphology and the fossil record. The probability of independent insertion at the same chromosomal address in multiple lineages is astronomically low for any single ERV; for hundreds, it is effectively zero under any hypothesis other than common descent.14
The GULO pseudogene provides a third independent line of evidence. Most mammals synthesize their own vitamin C using the enzyme L-gulonolactone oxidase, encoded by the GULO gene. Humans cannot synthesize vitamin C because the GULO gene is broken—it retains the sequence of a non-functional pseudogene carrying multiple inactivating mutations.9 Critically, chimpanzees, gorillas, and orangutans carry the same broken pseudogene with many of the same inactivating mutations in the same positions.9 The most parsimonious explanation is that the gene was broken once in a shared ancestor of all great apes and was inherited in its broken form by all descendants. Independent creation of separate species with the same non-functional gene carrying the same mutational history at the same genomic address has no plausible mechanism and no precedent.
Why the claims persist
Understanding why these objections persist despite the evidence requires distinguishing between scientific and cultural motivations. The scientific objections—that the fossil record is sparse, that individual specimens are ambiguous, that dating methods are unreliable—have been addressed repeatedly in the primary literature and in accessible science writing, yet the same objections recur in creationist publications with little acknowledgment of the responses. This pattern suggests that the objections serve a function other than genuine scientific inquiry.12
The morphological continuity of the hominin fossil record poses a specific theological challenge for traditions that hold that humanity was created separately from other animals and possesses a unique, non-biological origin. The genetic evidence is, if anything, more challenging, because it is harder to dismiss as ambiguous interpretation of fragmentary bones. The shared ERV insertions and chromosome 2 fusion are not matters of contested classification; they are specific, predicted molecular signatures that confirm common ancestry in a way that requires no subjective morphological judgment.7, 8, 14
Paleoanthropology and genomics operate by the same methods that produce every other result in modern biology and medicine. When creationist literature argues that the entire human fossil record has been systematically misread, or that molecular phylogenetics is unreliable, the argument requires a selective skepticism that is not applied to other conclusions drawn from the same methods. The evidence for human evolution from common ancestry is not a matter of a few contested specimens; it is the convergent result of fossil morphology, stratigraphy, comparative genomics, molecular phylogenetics, and biogeography, each developed independently and each pointing to the same conclusion.6, 12, 13
Summary of the evidence
The hominin fossil record documents a continuous morphological transition spanning more than 7 million years, from small-brained, partially arboreal forms near the base of the human lineage through obligate bipeds with ape-sized brains, into the genus Homo with its progressive expansion of the neocortex, reduction of facial anatomy, and development of complex technology.12, 13 Darwin’s 1871 prediction that human origins would prove African was confirmed by every subsequent fossil discovery. The genetic record—chromosome 2 fusion, shared ERV insertions, and the shared broken GULO gene—independently corroborates the pattern established by fossils and provides evidence of common ancestry that is not subject to interpretive ambiguity.7, 8, 9
The specific episodes cited as casting doubt on evolutionary science—Nebraska Man and Piltdown Man—were identified and corrected by scientists, not by critics of evolution. The “Lucy was just a chimp” claim is contradicted by every postcranial analysis of Australopithecus afarensis published in the peer-reviewed literature. The attempt to sort hominins into “ape or human” categories founders because the anatomical features that distinguish humans from apes change gradually and continuously across the fossil sequence, exactly as the hypothesis of common descent with modification predicts.3, 5, 6
References
Cloning and characterization of the human nonfunctional gene for L-gulono-gamma-lactone oxidase
Shared syntenic arrangements of retrovirus-like elements in the genomes of cats, cattle, and humans
Fossil footprints and the evolution of bipedalism: the functional significance of an early hominin foot