Overview
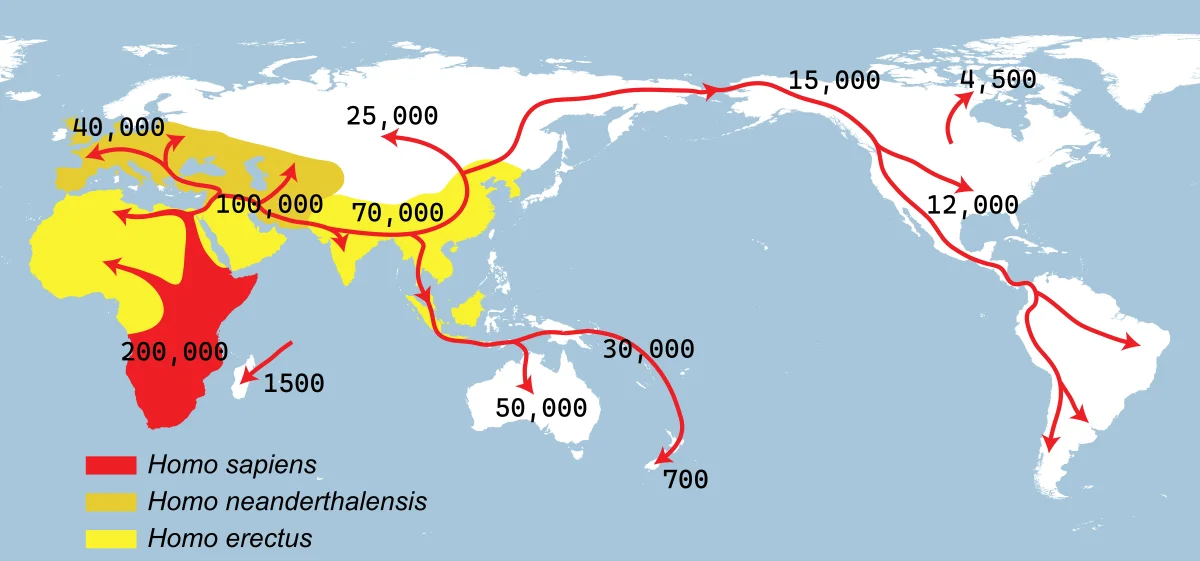
- After dispersing from Africa between approximately 70,000 and 50,000 years ago, Homo sapiens colonized the vast landmass of Eurasia through multiple routes and waves, reaching Australia and New Guinea by roughly 65,000 years ago, Southeast and East Asia by at least 45,000 years ago, and Europe by approximately 46,000 years ago, with earlier forays into the Levant and Arabia dating back more than 100,000 years.
- Ancient DNA from early Eurasian individuals, including the 45,000-year-old Ust'-Ishim man from Siberia and the Initial Upper Palaeolithic people of Bacho Kiro Cave in Bulgaria, reveals that the earliest modern human settlers of Eurasia carried recent Neanderthal ancestry and belonged to populations that contributed genetic legacy to present-day East Asians and Native Americans rather than to modern Europeans.
- The genetic structure of present-day Eurasian populations was shaped by successive waves of migration and admixture over tens of thousands of years, including the divergence of western and eastern Eurasian lineages by approximately 45,000 years ago, the expansion of Ancestral North Eurasians across Siberia, and massive Holocene population movements associated with the spread of farming from the Near East and the steppe pastoralist expansions of the Bronze Age.
The peopling of Eurasia is the story of how Homo sapiens, having evolved in Africa over the preceding 200,000 to 300,000 years, colonized the largest continuous landmass on Earth. This expansion was not a single event but a prolonged, multiphase process involving early forays into the Levant and Arabia more than 100,000 years ago, a major dispersal between roughly 70,000 and 50,000 years ago that ultimately gave rise to all non-African populations, and subsequent millennia of diversification, population replacement, and admixture that produced the genetic landscape of present-day Eurasia. Genomic analyses of both living and ancient DNA have shown that most non-African ancestry traces back to a single major Out of Africa expansion, though traces of earlier dispersals may survive in some populations, particularly in Oceania.1, 2
Along the way, modern humans encountered and interbred with archaic hominin populations that had been established in Eurasia for hundreds of thousands of years, principally Neanderthals in western Eurasia and Denisovans in eastern Eurasia. The genetic legacy of these encounters persists in every non-African human genome today. At the same time, the archaeological record documents a succession of technological and cultural innovations — the Initial Upper Palaeolithic, the Aurignacian, and subsequent traditions — that accompanied and perhaps facilitated the geographic expansion of modern humans across diverse and often challenging environments, from the tropical coastlines of Southeast Asia to the frozen steppe-tundra of Siberia.8, 9
{kind=link}
Early forays beyond Africa
The main Out of Africa dispersal was preceded by multiple earlier exits that brought Homo sapiens into the Levant and Arabia well before 70,000 years ago. A maxilla from Misliya Cave in Israel, dated to approximately 177,000 to 194,000 years ago, has been proposed as the earliest fossil evidence of modern humans outside Africa, though the dating and taxonomic assignment remain subjects of ongoing discussion.17 Fossils from the Skhul and Qafzeh cave sites in Israel, dated to approximately 90,000 to 120,000 years ago, demonstrate that anatomically modern humans maintained a presence in the Levant during warm interglacial phases. These populations appear to have coexisted with Neanderthals in the region before apparently retreating or being replaced, with Neanderthals reoccupying the Levant by around 70,000 years ago.14
{kind=link}
The Arabian Peninsula offers additional evidence of early dispersals. A Homo sapiens finger bone from the site of Al Wusta in Saudi Arabia, dated to approximately 85,000 years ago, demonstrates that modern humans had penetrated the interior of Arabia during a humid climatic phase when the Sahara and Arabian deserts were partially greened by increased monsoon rainfall.16 Climate modelling suggests that these "Green Arabia" episodes recurred at intervals determined by orbital precession cycles, opening corridors for human dispersal that closed again as aridity returned.22 Most of these early populations appear to have left little or no detectable genetic contribution to present-day non-African populations, suggesting that they were local expansions that did not persist through subsequent arid phases or that they were later swamped by the numerically larger main dispersal wave.14, 18
The pattern that emerges is one of repeated expansion and contraction, with Homo sapiens opportunistically colonizing areas beyond Africa during favourable climatic windows but failing to establish permanent populations until the conditions — demographic, technological, or environmental — aligned for a sustained expansion. Identifying what changed to make the main dispersal successful remains one of the central questions in human evolutionary studies.18
The main dispersal and its routes
The dispersal event that ultimately populated all of Eurasia, Oceania, and the Americas occurred between approximately 70,000 and 50,000 years ago. Genomic analyses consistently indicate that all non-African populations experienced a severe population bottleneck around this time, with effective population sizes falling to perhaps a few thousand individuals before expanding rapidly across new territories. This bottleneck is reflected in the markedly reduced genetic diversity of all non-African populations compared to African populations, a pattern that intensifies with increasing geographic distance from Africa in a phenomenon known as the serial founder effect.1
Two primary corridors have been proposed for the initial dispersal. The northern route through the Sinai Peninsula and the Levant would have brought modern humans into western Asia, from where they could disperse westward into Europe or eastward along the continental interior. The southern coastal route across the Bab el-Mandeb strait at the mouth of the Red Sea into the Arabian Peninsula and thence along the coastlines of South Asia toward Southeast Asia and Australia was popularized in the early 2000s as a mechanism to explain the rapid colonization of Australia by approximately 65,000 years ago.4, 14
Genomic data have complicated the simple two-route model. A 2016 study of 483 diverse genomes found that most non-African populations derive the great majority of their ancestry from a single dispersal event, but that Papuan populations carry a small percentage (~2%) of ancestry from an earlier dispersal wave distinct from the main expansion, suggesting that at least some early dispersers did contribute to living populations in Oceania.2 A companion genomic study of Aboriginal Australians estimated that their ancestors diverged from Eurasian populations approximately 51,000 to 72,000 years ago, consistent with a single major dispersal that subsequently branched rather than two entirely independent migrations.3 The current understanding is that the main dispersal was predominantly a single event, with minor contributions from earlier waves detectable in certain populations.
Colonization of Australia and Sahul
The colonization of Australia and New Guinea — then connected as the landmass Sahul during periods of low sea level — represents one of the earliest and most impressive achievements of the Eurasian dispersal. Reaching Sahul from mainland Southeast Asia required crossing the islands of Wallacea, a zone of deep water between the Asian and Australian continental shelves where even maximum sea-level lowering never created a continuous land bridge. Multiple sea crossings of at least 90 kilometres were necessary, implying that the colonizers possessed watercraft capable of open-ocean navigation.4
Excavations at the Madjedbebe rock shelter in northern Australia have yielded artefacts in primary depositional context dated by optically stimulated luminescence to approximately 65,000 years ago, making it the earliest securely dated site of human occupation on the continent. The associated toolkit includes grinding stones, ground ochres, reflective additives, and ground-edge hatchet heads, suggesting a technologically sophisticated founding population.4 Genomic analyses of Aboriginal Australian populations indicate a deep divergence from other non-African groups, followed by long-term genetic isolation. The study by Malaspinas and colleagues found that Papuan and Aboriginal Australian ancestors diversified from each other approximately 25,000 to 40,000 years ago, suggesting population structure within Sahul that predates the Holocene separation of Australia and New Guinea by rising sea levels.3
The implications of the Sahul colonization for understanding the broader Eurasian dispersal are significant. If modern humans reached northern Australia by 65,000 years ago, they must have traversed the coastlines of South and Southeast Asia considerably earlier, leaving a trail of now-submerged coastal sites that the archaeological record cannot yet document. The speed of the transit from Africa to Australia — potentially accomplished within a few thousand years — suggests rapid, directed movement along productive coastal environments rather than slow, diffusive population spread through continental interiors.4, 14
The earliest Eurasians in ancient DNA
Ancient DNA has transformed understanding of the earliest modern human populations in Eurasia. The oldest directly sequenced modern human genome is that of the Ust'-Ishim man, a Homo sapiens individual whose femur was recovered from the banks of the Irtysh River in western Siberia and radiocarbon-dated to approximately 45,000 years ago. His genome carries approximately the same proportion of Neanderthal DNA as present-day Eurasians (~2.3%), but the Neanderthal segments are substantially longer than those observed in living people, indicating that the admixture event had occurred only 7,000 to 13,000 years before he lived — roughly 50,000 to 60,000 years ago. Crucially, the Ust'-Ishim genome falls basal to the split between western and eastern Eurasian populations, suggesting he belonged to a population that lived before or during the initial divergence of these two major lineages.5

{kind=link}
The Initial Upper Palaeolithic (IUP) levels of Bacho Kiro Cave in Bulgaria have yielded Homo sapiens remains directly associated with IUP stone tools and radiocarbon-dated to approximately 43,000 to 46,000 years ago, representing one of the earliest securely dated modern human presences in Europe.6 Genome-wide sequencing of four of these individuals revealed that they carried 3 to 4 percent Neanderthal DNA, with the Neanderthal segments long enough to indicate a Neanderthal ancestor as recently as five to seven generations back. The finding that interbreeding with Neanderthals occurred repeatedly and recently in the family histories of these early Europeans was unexpected and suggests that admixture was common rather than exceptional when modern humans first entered Neanderthal territory.7
Equally surprising was the genetic affinity of the Bacho Kiro individuals. Rather than being ancestral to present-day Europeans, the oldest Bacho Kiro Cave individuals are more closely related to present-day East Asians and Native Americans. This finding implies that the earliest modern humans to reach Europe belonged to a population that either was subsequently replaced in the region or contributed its lasting genetic legacy primarily to populations further east.7 A similar pattern appears with the approximately 40,000-year-old Homo sapiens individual from Peștera cu Oase in Romania, who carried 6 to 9 percent Neanderthal DNA — indicating a Neanderthal ancestor only four to six generations back — but shows no detectable contribution to later European populations.21
Key early modern human genomes from Eurasia5, 6, 7, 21
| Individual | Location | Date (ka) | Neanderthal DNA | Closest living affinity |
|---|---|---|---|---|
| Ust'-Ishim | Western Siberia | ~45 | ~2.3% | Basal Eurasian (before E/W split) |
| Bacho Kiro (IUP) | Bulgaria | ~43–46 | ~3–4% | East Asians / Native Americans |
| Oase 1 | Romania | ~40 | ~6–9% | No detectable modern descendants |
Colonization and population turnover in Europe
The colonization of Europe by modern humans was a protracted process marked by multiple waves of settlement and repeated population turnover. The earliest arrivals, represented by the IUP toolkit and the Bacho Kiro Cave individuals, entered Europe via the Levant and the Danubian corridor by approximately 46,000 years ago. For several thousand years, modern humans and Neanderthals coexisted in different parts of the continent, with Neanderthals persisting in refugia on the Iberian Peninsula and elsewhere until approximately 40,000 years ago. By 35,000 years ago, Homo sapiens was the sole hominin species in Europe.6, 7
Analysis of 356 ancient hunter-gatherer genomes spanning the past 35,000 years has revealed that the genetic structure of European populations was far more dynamic than previously imagined. During the Last Glacial Maximum (LGM), approximately 26,500 to 19,000 years ago, human populations retreated into glacial refugia in southwestern Europe, the Italian peninsula, and the Balkans. The genetic composition of post-LGM Europe was dominated by the so-called Villabruna ancestry, associated with western European hunter-gatherers, which replaced or absorbed earlier genetic lineages across most of the continent as populations expanded from southwestern refugia after the ice retreated.11
The populations inhabiting Mesolithic Europe — the period between the end of the last Ice Age and the arrival of farming — were genetically distinct from the populations that followed. Analysis of ancient DNA has shown that present-day Europeans derive ancestry from at least three highly differentiated source populations: Western European Hunter-Gatherers (WHG), who contributed ancestry to all Europeans; Ancient North Eurasians (ANE), related to Upper Palaeolithic Siberians, who contributed to both Europeans and Near Easterners; and Early European Farmers (EEF), who were primarily of Near Eastern origin and arrived with the spread of agriculture beginning approximately 8,000 years ago.10 The arrival of farming peoples from Anatolia and the Levant transformed the genetic composition of Europe, with EEF ancestry largely replacing WHG ancestry across much of the continent.
A subsequent major genetic transformation occurred during the Bronze Age, approximately 5,000 to 4,000 years ago, when pastoralist populations from the Pontic-Caspian steppe expanded westward into Europe and eastward into Central and South Asia. These steppe pastoralists carried a distinctive mixture of Eastern European hunter-gatherer and Caucasus hunter-gatherer ancestry, and their expansion is associated with the spread of the Corded Ware and Yamnaya archaeological complexes and, most likely, the Indo-European language family. Steppe-related ancestry constitutes approximately 30 to 50 percent of the genome of present-day northern Europeans and is detectable across nearly all of Europe.10, 15
East Asia and Southeast Asia
The population history of East and Southeast Asia has been illuminated by a growing corpus of ancient DNA, though the record remains less dense than that available for Europe. Modern humans reached mainland East Asia by at least 40,000 years ago, as demonstrated by the Tianyuan individual from a cave near Beijing, whose genome shows clear East Asian affinity and Neanderthal ancestry comparable to that of other early Eurasians.13 The Japanese archipelago preserves evidence of human activity in the form of stone tools dating to approximately 38,000 years ago, and the earliest inhabitants of Japan — the Jomon people — represent a deeply divergent East Asian lineage that persisted in relative isolation for thousands of years.13
Ancient DNA from 166 East Asian individuals spanning the past 8,000 years has revealed a complex pattern of population structure and admixture. A deep genetic split separated northern and southern populations within East Asia, with Neolithic northern Chinese individuals being most closely related to modern-day Han Chinese and other East Asians, while ancient individuals from southern China showed closest affinity to modern-day Southeast Asians and Austronesian-speaking populations. Over the past several thousand years, a major southward expansion of northern East Asian ancestry progressively transformed the genetic landscape of southern China and Southeast Asia, reducing the genetic differentiation that had been much greater in the Neolithic period.20
The formation of present-day East Asian populations involved contributions from at least three major ancestry streams. Hunter-gatherers from Japan, the Amur River Basin, and peoples of Neolithic Taiwan and the Tibetan Plateau are linked by a deeply splitting lineage that probably reflects a coastal migration during the Late Pleistocene. Yellow River Basin farmers at approximately 3000 BCE spread Sino-Tibetan languages as their ancestry dispersed both to Tibet and to the Central Plain. Peoples from Taiwan between approximately 1300 BCE and 800 CE derived the majority of their ancestry from a lineage common among modern Austronesian, Tai-Kadai, and Austroasiatic speakers, likely descending from Yangtze River Valley farming populations.13 The genetic history of East Asia thus mirrors the pattern seen in Europe: an initial colonization by Pleistocene hunter-gatherers followed by massive population transformations driven by the expansion of farming peoples during the Holocene.
Siberia and Central Asia
The colonization of northern Eurasia — the vast interior of Siberia and Central Asia — presented formidable challenges. The subarctic and arctic environments of Siberia, with their extreme cold, short growing seasons, and sparse resources, were among the last regions of Eurasia to be permanently occupied by modern humans. Ancient DNA from 34 individuals spanning 31,000 to 600 years ago has revealed a complex population history involving at least three major migration events.12
The earliest known inhabitants of northeastern Siberia are represented by two milk teeth from the Yana Rhinoceros Horn Site (RHS), dated to approximately 31,600 years ago. The genomes of these Ancient North Siberians (ANS) revealed a previously unknown population that is distantly related to early western Eurasian hunter-gatherers but genetically distinct from any present-day population. The ANS people survived in extreme conditions at 71 degrees north latitude during the approach to the Last Glacial Maximum, subsisting on woolly mammoth, woolly rhinoceros, and bison.12
After the LGM, the genetic landscape of Siberia was transformed by the arrival of East Asian-related peoples, who admixed with ANS descendants to produce a population termed Ancient Palaeo-Siberians. This population is closely related to contemporary far-northeastern Siberian communities such as the Koryak and Itelmen, and also contributed significantly to the ancestry of Native Americans. The dual ancestry of Native Americans — a mixture of predominantly East Asian and Ancient North Eurasian components — can be traced to this admixture event, which occurred before or during the Beringian standstill period approximately 25,000 to 20,000 years ago.12
A third major migration wave, occurring during the Holocene, brought additional East Asian-related peoples into Siberia. These Neo-Siberians largely replaced the Palaeo-Siberian populations across much of the region and are the primary ancestors of many present-day Siberian peoples, including Yakuts, Evenks, and other Tungusic and Turkic-speaking groups.12 Central Asia experienced equally dramatic population transformations. Ancient DNA evidence from South and Central Asia has documented the expansion of steppe pastoralist ancestry into the region beginning approximately 4,000 years ago, coinciding with the decline of the Indus Valley Civilization and the likely spread of Indo-Iranian languages. The primary source of ancestry in modern South Asians is a prehistoric genetic gradient between people related to early hunter-gatherers of Iran and those of Southeast Asian affinity, with steppe pastoralist ancestry overlaid during the Bronze Age.15
Approximate dates of first modern human presence by region4, 5, 6, 12, 17
Archaic admixture across Eurasia
The dispersal of modern humans across Eurasia was accompanied by interbreeding with at least two archaic hominin populations: Neanderthals and Denisovans. The sequencing of the Neanderthal genome in 2010 revealed that all non-African modern humans carry approximately 1 to 4 percent Neanderthal DNA, while sub-Saharan Africans carry essentially none — a pattern consistent with admixture occurring after the Out of Africa dispersal but before the differentiation of major non-African populations.8
The geographic distribution and timing of Neanderthal admixture have been refined by ancient DNA. The Ust'-Ishim genome from 45,000 years ago carries Neanderthal segments long enough to indicate that admixture had occurred within the preceding 7,000 to 13,000 years, placing the primary mixing event at approximately 50,000 to 60,000 years ago, most likely in the Near East.5 The Bacho Kiro and Oase individuals demonstrate that additional, more local interbreeding events occurred as modern humans expanded westward into Europe, with some individuals having Neanderthal ancestors as recently as four to seven generations back.7, 21 The pattern suggests that admixture with Neanderthals was not a single event but occurred repeatedly along the advancing frontier of modern human expansion.
Denisovan ancestry shows a strikingly different geographic distribution. Present-day Melanesian and Aboriginal Australian populations carry approximately 4 to 6 percent Denisovan DNA, while mainland East Asian populations carry considerably less (~0.2%), and western Eurasians carry essentially none.9, 3 This distribution implies that the primary admixture between modern humans and Denisovans occurred in Southeast Asia or Near Oceania, consistent with the sparse Denisovan fossil record, which includes remains from Denisova Cave in the Altai Mountains of Siberia and a mandible from the Tibetan Plateau. Some introgressed archaic variants have been subject to positive natural selection in modern humans, including alleles affecting immune function and adaptation to specific environments.9, 8
The overall pattern is one of pervasive but low-level admixture: modern humans absorbed small amounts of archaic DNA as they expanded across Eurasia, with the specific archaic contribution varying by region depending on which archaic populations were encountered. Despite this interbreeding, the archaic populations themselves disappeared. Neanderthals vanished from the European record by approximately 40,000 years ago, and the latest Denisovan remains date to roughly 50,000 years ago. The mechanism of replacement likely involved a combination of competitive exclusion, demographic swamping by the larger modern human population, and possibly the effects of novel diseases.8, 19
Climate as a driver of dispersal
The timing and routes of human dispersal across Eurasia were strongly conditioned by climate. Climate modelling coupled with fossil and archaeological data has demonstrated that astronomically forced changes in temperature, rainfall, and terrestrial net primary production had a major impact on the distribution and viability of hominin habitats throughout the Pleistocene.19 The dispersal of Homo sapiens out of Africa and across Eurasia coincided with a period of fluctuating but generally declining global temperatures during Marine Isotope Stages 4 and 3 (roughly 71,000 to 29,000 years ago), punctuated by millennial-scale warm episodes (Dansgaard-Oeschger events) during which habitable zones expanded northward.
Coupled climate-migration modelling by Timmermann and Friedrich has shown that orbitally driven variations in African and Asian monsoon intensity created periodic windows of opportunity for human dispersal. During humid phases, the Sahara and Arabian deserts became traversable grasslands, while simultaneously the monsoon-fed river systems of South Asia provided corridors for eastward movement. During arid phases, these corridors closed, potentially isolating populations that had already dispersed and allowing them to diversify in geographic isolation.22
The Last Glacial Maximum posed the most severe environmental challenge to human populations in northern Eurasia. The expansion of the Scandinavian and Laurentide ice sheets, the descent of mountain glaciers, and the widespread development of permafrost rendered much of northern Europe and northern Asia uninhabitable. Human populations contracted into southern refugia — the Iberian Peninsula, the Italian and Balkan peninsulas, and possibly the Franco-Cantabrian region in southwestern Europe, as well as temperate zones in East Asia.11 The post-glacial recolonization of northern Eurasia from these refugia reshaped the genetic composition of the continent and set the stage for the Holocene population transformations that followed.
Holocene population transformations
The genetic landscape of present-day Eurasia was profoundly transformed during the Holocene (the past ~11,700 years) by three major demographic processes: the spread of agriculture from the Fertile Crescent and other centers of domestication, the expansion of pastoral nomads from the Eurasian steppe, and long-range maritime dispersals in Southeast Asia and the Pacific.
The agricultural expansion from the Near East into Europe, beginning approximately 8,500 years ago, brought Anatolian-derived farming populations into a continent previously inhabited exclusively by hunter-gatherers. Ancient DNA has shown that the transition to farming in most of Europe was accomplished primarily by the migration of farming peoples rather than by the adoption of farming by indigenous hunter-gatherers. Early European Farmers carried a genetic profile distinct from that of local Mesolithic hunter-gatherers, and the transition is marked by a sharp shift in ancestry composition across much of the continent.10 A parallel but independent expansion of farming peoples from the Fertile Crescent eastward into South Asia introduced Iranian-related ancestry that contributes substantially to the genomes of present-day South Asians.15
The steppe pastoralist expansion of the Bronze Age, beginning approximately 5,000 years ago, was equally transformative. Populations associated with the Yamnaya culture of the Pontic-Caspian steppe expanded both westward into Europe and eastward into Central and South Asia, carrying a distinctive genetic signature that combined Eastern European hunter-gatherer and Caucasus hunter-gatherer ancestry. In Europe, steppe ancestry replaced up to half of the previous Neolithic farmer ancestry in many regions, particularly in northern and central Europe. In South Asia, steppe-related ancestry is a defining component of Indo-European-speaking populations and is associated with the arrival of Vedic culture on the Indian subcontinent approximately 3,500 to 4,000 years ago.10, 15
In East and Southeast Asia, the expansion of farming peoples from the Yellow and Yangtze River valleys during the Neolithic drove analogous population transformations. The southward spread of Sino-Tibetan-speaking peoples from the Yellow River region and the maritime dispersal of Austronesian-speaking peoples from Taiwan reshaped the genetic composition of southern China, mainland Southeast Asia, and Island Southeast Asia over the past 5,000 years.13, 20 These Holocene migrations, driven by the demographic advantages conferred by food production, were often more consequential for the present-day genetic landscape of Eurasia than the original Pleistocene colonization events.
Synthesis and ongoing questions
The peopling of Eurasia emerges from the combined evidence of archaeology, paleoanthropology, ancient genomics, and climate science as a multi-layered process spanning more than 100,000 years. The earliest chapters involved repeated, tentative forays from Africa into the Levant and Arabia, most of which left little lasting genetic imprint. The major expansion, between roughly 70,000 and 50,000 years ago, gave rise to all non-African populations and was accomplished with remarkable speed: within 20,000 years or less, modern humans had colonized environments as diverse as the tropical forests of Wallacea, the temperate woodlands of Europe, the grasslands of Central Asia, and the tundra-steppe of Siberia.1, 18
Ancient DNA has revealed that the initial Pleistocene settlers of Eurasia were genetically distinct from the populations that inhabit the same regions today. The earliest Europeans, for example, were more closely related to present-day East Asians than to modern Europeans, and multiple waves of population replacement and admixture during and after the LGM completely reshaped the genetic structure of the continent.7, 11 Similar processes operated in East Asia, Siberia, and South Asia, where Holocene farming and pastoral expansions transformed the genetic landscape far more dramatically than the original colonization events.13, 15
Significant questions remain open. The role of early dispersals before 70,000 years ago is still poorly understood: beyond the ~2% trace detected in Papuan genomes, it remains unclear whether earlier waves contributed meaningfully to living populations anywhere in Eurasia.2 The archaeological record of the initial coastal dispersal is almost entirely submerged beneath post-glacial sea-level rise, leaving the route and pace of the migration poorly constrained. The mechanisms by which modern humans replaced Neanderthals and Denisovans — competitive superiority, demographic advantage, disease, climate disruption, or some combination — remain actively debated.8, 19 And the relationship between genetic populations and archaeological cultures, particularly during the critical transition from the Middle to the Upper Palaeolithic, is far from resolved.6
What the evidence has established with considerable confidence is that the peopling of Eurasia was neither a single migration nor a one-time event. It was a continuing process of expansion, contraction, replacement, and mixture that began more than 100,000 years ago and continues, in demographic terms, to the present day. The genomes of living Eurasian populations preserve a palimpsest of these successive layers, from the ancient Out of Africa bottleneck and the signatures of Neanderthal and Denisovan interbreeding to the massive population movements of the Holocene that gave the continent its current genetic structure.1, 10, 18
References
Ancient DNA indicates human population shifts and admixture in northern and southern China