Overview
- Genetic and archaeological evidence indicates that the ancestors of Native Americans diverged from East Asian and Ancient North Eurasian populations approximately 25,000 to 20,000 years ago, spent several thousand years isolated in Beringia during the Last Glacial Maximum, and then dispersed rapidly into the Americas by at least 16,000 years ago, with possible human presence as early as 23,000 years ago at White Sands, New Mexico.
- Ancient DNA from early American remains reveals that all Indigenous peoples of the Americas south of the Arctic descend primarily from a single founding population that subsequently diversified into northern and southern branches, with at least two later migration waves from Asia contributing ancestry to Arctic and Na-Dene-speaking populations.
- The route of initial entry likely followed the Pacific coast along a kelp highway of productive marine habitats rather than an interior ice-free corridor, as evidenced by the early appearance of sites like Monte Verde in southern Chile by 14,500 years ago and the chronological and genetic patterns of south-to-north diversification within the Americas.
The peopling of the Americas represents the last major continental colonization in human history. Beginning at some point during the Late Pleistocene, populations descended from northeastern Asian and Ancient North Eurasian lineages crossed the land bridge of Beringia into the Western Hemisphere and dispersed across two continents spanning from the Arctic to Tierra del Fuego. The timing, routes, and number of migration waves have been debated for more than a century, but since 2014 a convergence of ancient DNA evidence, improved radiocarbon chronologies, and new archaeological discoveries has transformed the field. The emerging picture is one of considerable complexity: an ancestral population that separated from East Asians by approximately 25,000 years ago, a period of isolation lasting several thousand years in Beringia during the Last Glacial Maximum, a rapid initial dispersal that reached the southern tip of South America within one to two millennia, and subsequent waves of migration that contributed additional Asian ancestry to Arctic and subarctic populations.1, 2
The question of when humans first entered the Americas has proved particularly contentious. For much of the twentieth century, the Clovis culture, dating to approximately 13,000 calendar years before present, was considered the earliest unambiguous evidence of human habitation. The discovery of pre-Clovis sites, culminating in the 2021 report of human footprints at White Sands, New Mexico, dating to approximately 23,000–21,000 years ago, has pushed the timeline substantially earlier and reopened fundamental questions about the relationship between archaeological visibility and actual human presence.10, 11
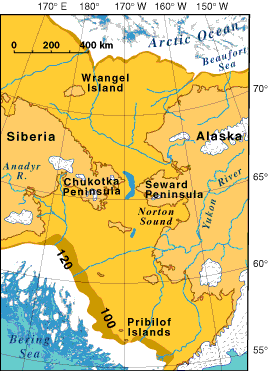
Beringia and the land bridge
The entry of humans into the Americas was made possible by a geological and climatic circumstance: the periodic emergence of a vast land connection between northeastern Asia and northwestern North America during glacial periods when global sea levels dropped by as much as 120 metres below present levels. This land bridge, known as Beringia, was not a narrow isthmus but a broad subcontinent, at its maximum extent stretching approximately 1,600 kilometres from north to south and encompassing what is now the shallow floor of the Bering and Chukchi seas. During the Last Glacial Maximum (LGM), roughly 26,500 to 19,000 years ago, Beringia was an exposed, treeless steppe-tundra supporting populations of mammoth, bison, horse, and caribou — and, the evidence now suggests, human hunters as well.1, 14
{kind=link}
The Beringian standstill hypothesis, formulated by Erika Tamm, Toomas Kivisild, and colleagues in 2007, proposed that the ancestors of Native Americans did not pass rapidly through Beringia but instead occupied it for an extended period — perhaps 5,000 to 15,000 years — during which their mitochondrial DNA lineages diverged from Asian sister-clades and accumulated the mutations that characterize the founding haplogroups of the Americas (A, B, C, D, and X).1 The hypothesis was based on the observation that all major Native American mitochondrial lineages coalesce to dates substantially older than the earliest archaeological evidence of human presence south of the ice sheets, implying that the founding population must have existed as a genetically distinct entity well before dispersing into the continental interior. Whole-genome analyses have subsequently confirmed this model, placing the separation of the Ancestral Native American lineage from East Asian populations at approximately 25,000 to 20,000 years ago and estimating the Beringian isolation period at roughly 2,400 to 9,000 years.3, 5
The ancestral composition of the founding population has been clarified by the sequencing of the Mal'ta genome, a 24,000-year-old boy from south-central Siberia whose DNA revealed a population — termed Ancient North Eurasians (ANE) — that contributed approximately 35% of the ancestry of Native Americans, with the remaining approximately 65% derived from an East Asian source population.19 This dual ancestry, East Asian and ANE, is a defining genetic signature of all Native American populations and distinguishes them from any single present-day Asian group. The admixture event that produced this combination is estimated to have occurred before or during the Beringian standstill, between approximately 25,000 and 20,000 years ago.3, 19
Routes of entry into the Americas
Two primary routes have been proposed by which humans could have moved from Beringia into the Americas south of the continental ice sheets: an interior corridor between the Laurentide and Cordilleran ice sheets, and a Pacific coastal route along the margin of the continent.
The ice-free corridor hypothesis dominated thinking for most of the twentieth century. As the massive ice sheets that covered most of Canada during the LGM began to retreat, a narrow passage opened between the Laurentide ice sheet (centred on Hudson Bay) and the Cordilleran ice sheet (covering the Canadian Rockies and the Pacific coastal ranges). This corridor would have provided a terrestrial route from interior Alaska southward into the northern Great Plains. However, environmental DNA analysis of lake sediment cores from the corridor region has demonstrated that the corridor did not become biologically viable — that is, capable of supporting the plant and animal communities necessary to sustain human migration — until approximately 12,600 years ago, several centuries after the earliest well-dated archaeological sites south of the ice sheets.18 This chronological mismatch means that while the ice-free corridor may have served as a route for later migrations, it was not available for the initial entry of people into the Americas.
The Pacific coastal route, by contrast, may have been accessible significantly earlier. The kelp highway hypothesis, articulated by Jon Erlandson and colleagues in 2007, proposes that Late Pleistocene maritime peoples could have migrated southward along the Pacific rim by exploiting the continuous belt of highly productive kelp forest ecosystems that stretches from Japan through the Aleutian Islands and down the west coast of North America to Baja California.13 Kelp forests support dense populations of shellfish, fish, marine mammals, and edible seaweeds, and would have provided a reliable subsistence base for coastal peoples equipped with watercraft. Glaciological evidence suggests that portions of the outer coast of southeastern Alaska and British Columbia were deglaciated by 16,000 to 15,000 years ago, and that offshore islands and exposed sections of the continental shelf would have offered refugia even earlier.13, 14
The principal difficulty with testing the coastal route hypothesis is that Pleistocene shorelines are now submerged beneath as much as 120 metres of water, and the archaeological sites of any early coastal migrants would have been destroyed or buried by rising sea levels at the end of the ice age. Nevertheless, the circumstantial evidence is substantial: the earliest sites south of the ice sheets, including Monte Verde in Chile and several sites along the Pacific coast of South America, are either coastal or contain marine resources, and the rapidity with which humans appear to have reached southern South America is most parsimoniously explained by movement along the coast rather than through the continental interior.12, 14
Archaeological evidence for early habitation
The archaeological record of the earliest Americans has undergone a revolution in the past two decades. For much of the twentieth century, the Clovis culture, characterised by its distinctive fluted bifacial projectile points and dated to approximately 13,000 to 12,700 calendar years before present, was regarded as representing the first inhabitants of the Americas. The "Clovis First" model held that big-game hunters crossed the ice-free corridor and rapidly colonised the continent, their large, finely crafted points representing the technological signature of the founding population.15 Clovis sites have been identified across much of North America, and the associated toolkit is remarkably uniform across a vast geographic range, suggesting rapid dispersal by a single, technologically coherent population.
The Clovis First model collapsed under the weight of accumulating evidence from sites that predate the Clovis horizon. The most important of these is Monte Verde, an open-air settlement on the banks of Chinchihuapi Creek in southern Chile, excavated by Tom Dillehay and colleagues beginning in the 1970s. The upper occupation layer, Monte Verde II, has been radiocarbon-dated to approximately 14,500 calendar years before present and contains preserved wooden foundations of a long residential structure, clay-lined hearths, stone and bone tools, and remains of at least sixty species of edible and medicinal plants, including nine species of seaweed transported from the coast some 90 kilometres away.12 The acceptance of Monte Verde as a pre-Clovis site by a visiting committee of archaeologists in 1997 was a watershed moment that opened the field to the possibility of earlier migrations.
Additional pre-Clovis sites have since gained acceptance, including the Paisley Caves in Oregon (human coprolites dated to approximately 14,300 years ago), the Buttermilk Creek complex at the Debra L. Friedkin site in Texas (artifacts dated to approximately 15,500 years ago), and the Cooper's Ferry site in Idaho (stemmed projectile points dated to approximately 16,000 years ago).14 The most provocative discovery has been the White Sands footprints in New Mexico. In 2021, Matthew Bennett, Jeffrey Pigati, and colleagues reported the discovery of in situ human footprints stratigraphically located between layers of sediment containing seeds of the aquatic plant Ruppia cirrhosa that were radiocarbon-dated to approximately 23,000 to 21,000 years ago — squarely within the Last Glacial Maximum.10 Initial scepticism focused on the possibility that the Ruppia seeds might yield anomalously old dates due to a hard-water effect, in which aquatic plants incorporate ancient carbon from their underwater environment. However, a 2023 study by Pigati and colleagues redated the site using radiocarbon measurements of conifer pollen extracted from the same stratigraphic layers and optically stimulated luminescence dating of quartz grains, obtaining ages of 20,700 to 22,400 years ago — closely matching the original seed-based dates and substantially strengthening the case for a human presence in the Americas during the LGM.11
The Clovis culture and the Anzick-1 genome
Although Clovis is no longer regarded as the earliest cultural manifestation in the Americas, it remains one of the most important. The Clovis technocomplex, defined by its large, fluted lanceolate points and dating to approximately 13,000 to 12,700 calendar years before present, represents the earliest widely distributed archaeological tradition across North America. Clovis points have been found from Nova Scotia to Mexico, frequently in association with the remains of large Pleistocene megafauna including mammoth, mastodon, and bison, leading to the traditional interpretation of Clovis people as specialised big-game hunters, though recent analyses suggest a broader subsistence strategy that included plant foods and smaller game.15

.jpg){kind=link}
Refined radiocarbon dating of Clovis sites by Michael Waters and Thomas Stafford in 2007 compressed the Clovis temporal range to a span of only approximately 200 to 350 years, suggesting a remarkably rapid cultural florescence and dispersal rather than a long, gradual colonisation process.15 The brevity of the Clovis window implies that Clovis technology either developed very quickly in situ within the Americas or was brought as a fully formed toolkit by a population that expanded rapidly across the continent.
The relationship between Clovis people and contemporary Native Americans was established definitively in 2014 with the sequencing of the genome of Anzick-1, a male infant interred with Clovis artifacts at a site in western Montana dated to approximately 12,600 calendar years before present. Rasmussen and colleagues found that the Anzick-1 genome is more closely related to all present-day Indigenous American populations than to any other group in the world, directly refuting hypotheses that Clovis people were related to Upper Palaeolithic Europeans (the so-called Solutrean hypothesis) or represented a population replaced by later arrivals.6 The Anzick-1 genome also confirmed the dual ancestry pattern (East Asian plus Ancient North Eurasian) characteristic of all Native Americans, with the ANE component matching the Mal'ta boy from Siberia. The gene flow from ANE into Native American ancestors must therefore have occurred before approximately 12,600 years ago, consistent with the Beringian standstill model.6, 19
Ancient DNA and population structure
The application of whole-genome sequencing to ancient American remains has revealed a population history of unexpected complexity. The founding population of the Americas diverged from East Asian source populations approximately 25,000 to 20,000 years ago and subsequently split into at least two major branches: a northern branch that includes most present-day northern North American populations, and a southern branch ancestral to most Central and South American Indigenous peoples as well as some North American groups.3, 5
A key discovery came from the Upward Sun River site in interior Alaska, where the remains of two infants (USR1 and USR2), dated to approximately 11,500 years ago, were recovered from a residential hearth. Sequencing of the USR1 genome by Moreno-Mayar and colleagues in 2018 revealed a previously unknown population, termed Ancient Beringians, that diverged from the common ancestor of all other Native Americans approximately 22,000 to 18,000 years ago but remained in Alaska while the remainder of the founding population dispersed southward.3 The Ancient Beringians represent a basal branch of the Native American family tree that has no identifiable living descendants, as their genetic signature was subsequently absorbed by later-arriving populations. The divergence date of the Ancient Beringians is consistent with the Beringian standstill model and places the ancestral population in eastern Beringia during the LGM.3
The northern and southern branches of Native Americans diverged from each other approximately 17,500 to 14,600 years ago, a date that postdates the opening of coastal migration routes but predates the formation of the ice-free corridor as a biologically viable passage.3, 18 This timing supports the interpretation that the initial dispersal proceeded along the Pacific coast and that the two branches separated as some populations moved into the interior of North America while others continued southward. The southern branch diversified rapidly upon entering Central and South America, with multiple parallel expansions documented by ancient genomes spanning the past 11,000 years from sites in Belize, Brazil, the Central Andes, and the Southern Cone.8
Estimated divergence dates in Native American population history2, 3, 5
| Divergence event | Estimated date (ka) | Method |
|---|---|---|
| Ancestral Native Americans from East Asians | ~25–20 | Whole-genome coalescence |
| ANE admixture into founding population | ~25–20 | Admixture graph modelling |
| Ancient Beringians from other Native Americans | ~22–18 | Whole-genome coalescence (USR1) |
| Northern and southern Native American branches | ~17.5–14.6 | Whole-genome coalescence |
| South American diversification | ~14–11 | Ancient genome time transects |
Multiple waves of migration
While the initial peopling of the Americas was accomplished by a single founding population, subsequent millennia saw additional waves of migration from Asia that contributed genetic ancestry to specific populations. David Reich and colleagues, analysing genome-wide data from 52 Native American and 17 Siberian groups, identified at least three streams of Asian gene flow into the Americas.7 The first and by far the largest, which they termed the "First American" stream, accounts for the great majority of ancestry in all Native American populations from Canada to the tip of South America. The second stream contributed approximately half the ancestry of Eskimo-Aleut speakers in the Arctic, representing a migration that occurred approximately 4,500 years ago and is associated with the Palaeo-Eskimo archaeological tradition. The third stream contributed roughly one-tenth of the ancestry of Na-Dene-speaking populations, including Chipewyan and other Athabaskan groups.7
Ancient DNA from Palaeo-Eskimo sites in Greenland and the Canadian Arctic has confirmed the second migration wave. The genome of the approximately 4,000-year-old Saqqaq individual from Greenland showed close affinity with present-day Chukchi and other northeastern Siberian populations rather than with modern Inuit, indicating a migration from Siberia that was partially independent of the earlier First American movement.2 The Saqqaq/Dorset Palaeo-Eskimo tradition persisted for roughly 4,000 years before being replaced or absorbed by the Thule culture, the direct ancestors of modern Inuit, who expanded from Alaska eastward across the Arctic approximately 1,000 years ago in a third identifiable migration event.2, 7
Within the Americas, post-initial-colonisation gene flow events have also been documented. Posth and colleagues identified at least two previously unrecognised streams of gene flow between North and South America, including one that affected the Central Andes by approximately 4,200 years ago, replacing much of the earlier local ancestry with ancestry related to Mesoamerican populations.8 These later movements document the ongoing dynamism of American population history long after the initial colonisation.
The Australasian ancestry signal
One of the most unexpected findings from genomic studies of Native Americans is the detection of a statistical affinity between certain Amazonian populations and present-day Australasian peoples — Aboriginal Australians, Melanesians, and the Andamanese. In 2015, Pontus Skoglund and colleagues reported that three Amazonian groups — the Surui, Karitiana, and Xavante — share more genetic variants with Australasians than do other Native American populations, a pattern they attributed to a ghost population they designated "Population Y" (from the Tupi-Guarani word Ypykuera, meaning "ancestor").9 Population Y, they argued, reached the Americas either before or around the same time as the main First American migration, more than 15,000 years ago, and contributed a small but detectable proportion of ancestry to some South American groups while leaving no apparent trace in North American populations.
The Australasian signal has been confirmed by multiple independent studies, including analyses of ancient genomes from Brazil dating to approximately 10,000 years ago.4, 8 Moreno-Mayar and colleagues, sequencing 15 ancient American genomes spanning from Alaska to Patagonia, found that the Australasian affinity was detectable in some ancient South American individuals but absent from all ancient and modern North American populations they examined, strengthening the case for a separate early migration event that contributed to South American but not North American populations.4 The route by which Population Y reached South America remains unknown. Possibilities include an early coastal migration that preceded the main First American expansion, a trans-Pacific route, or descent from a structured Beringian population in which different subgroups carried different proportions of Australasian-related ancestry. The signal is too faint and geographically restricted to resolve with current data, and it remains one of the most intriguing unsolved puzzles in the genetics of the Americas.2, 9
Speed of colonisation and demographic expansion
One of the most striking features of the peopling of the Americas is the speed with which it occurred. If the initial entry happened approximately 16,000 years ago via the Pacific coast, then the presence of humans at Monte Verde in southern Chile by approximately 14,500 years ago implies that populations traversed more than 14,000 kilometres of coastline and reached the southern cone of South America within roughly 1,500 years.12, 14 This translates to an average dispersal rate on the order of 10 kilometres per year, a pace consistent with the movement of small, mobile groups of coastal hunter-gatherers exploiting a linear belt of marine resources.
Genetic evidence corroborates this rapid expansion. Analyses of ancient mitochondrial genomes have shown that the founding mitochondrial lineages of the Americas diversified in a burst-like pattern, with multiple major branches emerging within a narrow time window of approximately 16,000 to 13,000 years ago.17 This demographic signature — a "star-like" phylogeny in which many lineages radiate from a single point of origin within a short interval — is characteristic of rapid population expansion into previously unoccupied territory. The pattern is mirrored in Y-chromosome data and in the distribution of autosomal haplotype blocks.
Approximate dates of earliest evidence per region2, 10, 12, 14
The demographic expansion that accompanied the initial colonisation was enormous. From a founding population estimated at perhaps a few thousand individuals, the population of the Americas grew to tens of millions by the time of European contact, filling ecological niches from tropical rainforests to Arctic tundra, high-altitude plateaux to temperate grasslands. This population growth involved not just geographic expansion but rapid local adaptation to diverse environments, with genetic evidence documenting selection for high-altitude tolerance in Andean populations, dietary adaptations in Arctic peoples, and variation in immune function across the continent.2, 16
Arrival of humans and megafaunal extinction
The entry of humans into the Americas coincided closely with the extinction of the continent's megafauna. Between approximately 15,000 and 10,000 years ago, the Americas lost at least 35 genera of large mammals, including mammoths, mastodons, ground sloths, sabre-toothed cats, American horses, camels, and glyptodonts. The causes of this extinction remain debated, with three primary hypotheses: direct human predation ("overkill"), climate change associated with the end of the Pleistocene, and a synergistic interaction between the two.14
The overkill hypothesis, originally articulated by Paul Martin in the 1960s, argues that naive megafaunal populations that had never encountered human predators were rapidly hunted to extinction by skilled, technologically equipped hunters expanding into the continent. The temporal correlation between human arrival and extinction is striking: the megafaunal losses in the Americas occurred thousands of years after similar-magnitude climate changes that did not produce comparable extinctions, and the chronology of megafaunal disappearance generally tracks the southward advance of human populations.14 Clovis sites in particular frequently contain the remains of mammoths and other large animals in clear association with hunting tools, providing direct evidence that early Americans hunted megafauna.
The climate-change hypothesis points to the dramatic environmental reorganisation that occurred at the end of the Pleistocene, as warming temperatures altered vegetation patterns, fragmented habitats, and disrupted the ecological relationships on which large-bodied herbivores depended. Many of the extinct species had survived multiple previous glacial-interglacial cycles, however, a point that advocates of the overkill hypothesis emphasise. The most widely accepted current view holds that the extinction resulted from a combination of both factors: climate-driven habitat fragmentation reduced megafaunal populations to vulnerable levels, and the added pressure of human hunting pushed them past the point of recovery.14 Whatever the relative contributions of the two forces, the loss of the megafauna profoundly shaped the ecology of the Americas and the subsistence strategies of the human populations that remained.
Synthesis and open questions
The current understanding of the peopling of the Americas can be summarised in a multi-stage model. First, a source population in northeastern Asia admixed with Ancient North Eurasians approximately 25,000 to 20,000 years ago and moved into Beringia, where it was isolated for several thousand years during the Last Glacial Maximum. Second, this Beringian population began dispersing southward by at least 16,000 years ago, most likely along the Pacific coast, reaching southern Chile by approximately 14,500 years ago. Third, the initial founding population diverged into northern and southern branches, with subsequent internal diversification producing the rich mosaic of Indigenous American peoples present at the time of European contact. Fourth, at least two additional migration waves from Asia introduced Palaeo-Eskimo and Thule ancestry to the Arctic, and Na-Dene-related gene flow contributed to subarctic populations.2, 3, 7
Significant questions remain open. The White Sands footprints, if their dating is confirmed by additional studies, would require a fundamental reassessment of the timeline, pushing human presence in the Americas back to approximately 23,000 years ago and implying that an early population was present during the LGM but either did not survive or left no detectable genetic legacy in later populations.10, 11 The nature and route of the Australasian-related Population Y that contributed ancestry to some South American populations remains entirely unresolved.9 The question of whether the ice-free corridor served as a secondary route for north-south migration after it became biologically viable around 12,600 years ago, even if it was not the initial entry route, awaits further archaeological and genetic evidence.18
The relationship between archaeological cultures and genetic populations is also far from straightforward. The Clovis technology appears across a vast area in a remarkably short time span, but whether it represents the spread of people, the spread of ideas, or some combination of both cannot be resolved by archaeology alone and awaits additional ancient genomes from pre-Clovis and Clovis-age individuals.6, 15 Similarly, the processes by which later population movements — including the Mesoamerican-related expansion into South America documented by Posth and colleagues — reshaped the genetic landscape of the Americas are only beginning to be understood.8
What the convergence of genetic, archaeological, and environmental evidence has established beyond reasonable doubt is that the peopling of the Americas was not a single event but a process extending over thousands of years, involving multiple populations, multiple routes, and continuous adaptation to some of the most diverse environments on Earth. The ancestors of today's Indigenous American peoples accomplished one of the most remarkable feats of human dispersal in history, colonising two continents spanning 14,000 kilometres of latitude within a few millennia and developing the cultural, linguistic, and biological diversity that characterised the pre-contact Americas.2, 14
References
Terminal Pleistocene Alaskan genome reveals first founding population of Native Americans
Genomic evidence for the Pleistocene and recent population history of Native Americans
Independent age estimates resolve the controversy of ancient human footprints at White Sands
The kelp highway hypothesis: marine ecology, the coastal migration theory, and the peopling of the Americas
Ancient mitochondrial DNA provides high-resolution time scale of the peopling of the Americas