Overview
- Dunkleosteus terrelli was a giant arthrodire placoderm fish that inhabited Late Devonian seas approximately 358–382 million years ago, making it one of the largest and most formidable vertebrate predators of the Paleozoic era, with an armored head shield composed of interlocking dermal bone plates up to 5 centimeters thick.
- Biomechanical analyses of Dunkleosteus jaw mechanics indicate it could generate bite forces of approximately 6,000 newtons at the jaw tip and over 7,400 newtons at the rear dental plates, rivaling the bite force of large crocodilians and making it among the most powerful biters in the history of vertebrate life.
- Recent body size reassessments have significantly revised estimates of Dunkleosteus total length downward from the frequently cited 6–10 meters to approximately 3.4–4.1 meters, based on new comparisons with complete arthrodire body plans rather than extrapolation from great white sharks.
Dunkleosteus is an extinct genus of large placoderm fish belonging to the order Arthrodira that lived during the Late Devonian period, approximately 358 to 382 million years ago.1, 8 The best-known and largest species, Dunkleosteus terrelli, is represented primarily by fossils from the Cleveland Shale of Ohio, a dark, organic-rich marine shale deposited in an epicontinental sea that covered much of what is now the eastern United States during the Famennian stage of the Late Devonian.1, 9 The genus was named in 1956 in honor of David Dunkle, a curator of vertebrate paleontology at the Cleveland Museum of Natural History, who spent decades studying the Devonian fishes of the Cleveland Shale.1 As a heavily armored apex predator with powerful jaw mechanisms, Dunkleosteus has become one of the most iconic animals of the Paleozoic era and a symbol of the extraordinary vertebrate diversity that characterized the Devonian, often called the "Age of Fishes."7, 8
{kind=link}
Anatomy and the armored head shield
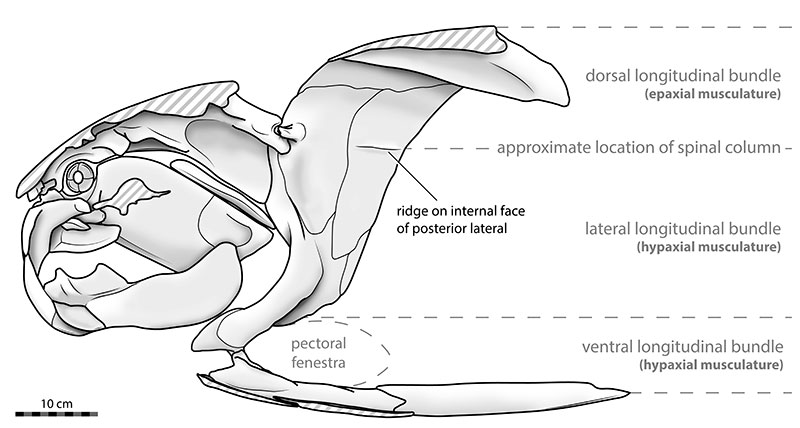
Like all arthrodire placoderms, Dunkleosteus possessed a body divided into two principal regions encased in dermal bone: a head shield (cranial shield) covering the skull and gill region, and a trunk shield protecting the anterior portion of the body.1, 8 The two shields were connected by a mobile joint at the back of the head, the craniothoracic joint, which allowed the head to be tilted upward during jaw opening, a mechanism unique to arthrodires that significantly increased the gape of the mouth.1, 2 The head shield of D. terrelli is composed of interlocking dermal bone plates including the central, nuchal, paranuchal, postorbital, preorbital, and rostral plates, which together form a rigid, helmet-like structure that was originally up to approximately 5 centimeters thick in large individuals.1, 15
The jaws of Dunkleosteus are among its most remarkable features. Unlike the jaws of modern bony fishes and sharks, which bear separate teeth attached to the jawbones, the jaws of Dunkleosteus consisted of sharpened bony plates, the inferognathal (lower jaw) and supragnathal (upper jaw) elements, whose edges were honed into self-sharpening blades by the shearing action of the opposing plates against each other.1, 2 The anterior portions of these plates came to sharp, fang-like points that would have functioned for piercing prey, while the posterior portions formed broad, blade-like surfaces suited for cutting and crushing.1, 3 This dual-function jaw morphology suggests that Dunkleosteus was capable of handling a wide range of prey items, from soft-bodied organisms to heavily armored fish.2, 10
The posterior portion of the body, behind the trunk shield, is known almost entirely from inference rather than direct fossil evidence. Because placoderm trunk skeletons were composed of cartilage rather than bone, they rarely fossilized, and no complete body of D. terrelli has been recovered.5, 8 For over a century, reconstructions of the full animal were based on scaling from the body proportions of modern great white sharks, which led to widely reproduced estimates of 6 to 10 meters in total length.8 However, more recent studies have challenged these estimates substantially, as discussed below.5
Bite force and feeding mechanics
Philip Anderson and Mark Westneat published a series of influential biomechanical analyses of Dunkleosteus jaw mechanics in 2007 and 2009, using lever-arm models and computational simulations based on the detailed anatomy of the Cleveland Museum skull.2, 3 Their 2007 study modeled the jaw adductor musculature and the craniothoracic joint mechanism, concluding that the unique four-bar linkage system of the arthrodire skull allowed Dunkleosteus to open its jaws extremely rapidly, completing a full gape cycle in approximately 20 milliseconds, creating a strong suction force that would have helped to draw prey into the mouth.2
The 2009 study focused on bite force estimation and calculated that D. terrelli could generate bite forces of approximately 6,000 newtons (N) at the anterior tip of the jaw and over 7,400 N at the posterior bladed region, where the mechanical advantage of the jaw lever system was greatest.3 These values are comparable to the bite forces estimated for large modern saltwater crocodiles and significantly exceed those of any living fish species.3 Anderson and Westneat noted that the combination of high bite force at the rear dental plates with a rapid, suction-generating jaw opening made Dunkleosteus a formidable predator, capable of both capturing fast-moving prey through suction and processing heavily armored prey through powerful crushing bites.2, 3
Evidence for the predatory capabilities of Dunkleosteus extends beyond biomechanical models. Robert Carr and Gene Jackson described a specimen of a smaller arthrodire, Holonema, that preserved apparent bite marks matching the jaw morphology of Dunkleosteus, providing direct evidence of predator-prey interactions in the Cleveland Shale ecosystem.10 Several Dunkleosteus specimens also preserve clusters of semi-digested fish bones within the body cavity, interpreted as regurgitated or partially processed gut contents, indicating that Dunkleosteus consumed other large fish, including smaller placoderms.1, 10
Body size reassessment
For decades, Dunkleosteus terrelli was depicted in museum exhibits and popular media as a colossal fish reaching 6 to 10 meters in total length, a figure derived by scaling the known head and trunk shield dimensions to the body proportions of a great white shark (Carcharodon carcharias).8 This approach assumed that the post-thoracic body of Dunkleosteus was proportioned similarly to that of a large macropredatory shark, but no direct fossil evidence supported this assumption.5
In 2023, Russell Engelman published a comprehensive reassessment of Dunkleosteus body size using comparisons with complete arthrodire specimens from other genera, rather than with sharks.5 Engelman argued that arthrodires had proportionally much larger heads relative to their total body length than sharks do, and that using shark proportions as a scaling model therefore systematically overestimated the total length of Dunkleosteus.5 Using a dataset of complete arthrodire body fossils and multiple regression approaches, Engelman estimated that the largest known D. terrelli specimens had total body lengths of approximately 3.4 to 4.1 meters, roughly one-third to one-half of the traditional estimates.5 This revised body plan depicts Dunkleosteus as a stocky, heavily built predator with a very large head relative to its body, more tuna-like or grouper-like in overall proportions than shark-like.5
Engelman's revision has been supported by additional work by Jobbins and colleagues, who compared body size estimation methods across multiple placoderm lineages and concluded that traditional estimates for large arthrodires were indeed inflated by inappropriate scaling models.14 Even at the revised lengths, Dunkleosteus remains one of the largest known Devonian vertebrates and was almost certainly an apex predator in its ecosystem, but the image of a 10-meter leviathan has given way to a more compact but no less formidable animal.5, 14
Ecology and the Cleveland Shale
The vast majority of D. terrelli specimens come from the Cleveland Shale, a geological unit of the Ohio Shale sequence exposed along the south shore of Lake Erie in northeastern Ohio, with particularly rich fossil localities in the Rocky River valley and the Big Creek valley near Cleveland.9 The Cleveland Shale was deposited in a shallow epicontinental sea during the Famennian stage of the Late Devonian, approximately 358 to 372 million years ago, under conditions of low-oxygen bottom waters that favored exceptional preservation of organic material, including the phosphatic bones and dermal plates of placoderms.9, 13
The Cleveland Shale fauna represents a complex marine ecosystem dominated by placoderms. In addition to Dunkleosteus, the formation has yielded fossils of the giant filter-feeding arthrodire Titanichthys, which reached comparable head shield dimensions but possessed weak, toothless jaws suited for planktivorous feeding rather than predation.16 Smaller arthrodires including Holonema and Heintzichthys, various ptyctodonts and antiarchs, early sharks such as Cladoselache, and acanthodians round out the vertebrate fauna.9, 10 The co-occurrence of Dunkleosteus and Titanichthys in the same deposits illustrates the ecological diversity of large arthrodires, which occupied trophic roles ranging from apex predation to suspension feeding.16
Dunkleosteus is not restricted to the Cleveland Shale. Specimens attributed to the genus or closely related forms have been reported from Late Devonian deposits in Belgium, Morocco, Poland, and possibly Australia, indicating a broad geographic distribution across Late Devonian marine environments.8 However, the Cleveland Shale remains by far the most prolific source of well-preserved material, and the detailed anatomy of the genus is known primarily from Ohio specimens curated at the Cleveland Museum of Natural History.1, 9
Evolutionary significance and extinction
Dunkleosteus and the arthrodire placoderms to which it belongs represent a major chapter in the evolutionary history of jawed vertebrates. Placoderms were the first jawed vertebrates to dominate marine ecosystems, and they radiated explosively during the Devonian to fill ecological roles ranging from benthic detritivores to pelagic filter feeders to apex predators.7, 8 The jaw structures of placoderms, composed of dermal bone plates rather than separate teeth, represent an early and distinct solution to the biomechanical challenges of prey capture and processing, one that was ultimately superseded by the tooth-bearing jaws of osteichthyans (bony fishes) and chondrichthyans (sharks and rays) but was highly effective for over 50 million years.6, 7, 12
The phylogenetic relationships of placoderms have been debated extensively. Traditional classifications placed them as a monophyletic group, the class Placodermi, basal to the divergence of osteichthyans and chondrichthyans.8 More recent cladistic analyses by Martin Brazeau, Matt Friedman, and others have suggested that placoderms may be paraphyletic, with some lineages more closely related to the ancestors of modern jawed vertebrates than others, which would make the placoderm body plan ancestral to all modern gnathostomes rather than a separate evolutionary dead end.7 Under this interpretation, studying Dunkleosteus and its relatives is essential for understanding the origin and early diversification of the jaws, teeth, and skeletal structures that characterize all living jawed vertebrates.6, 7
Dunkleosteus and all other placoderms went extinct at or near the end of the Devonian period, during a series of environmental crises collectively known as the Late Devonian extinction events.13 The Frasnian-Famennian boundary event, approximately 372 million years ago, eliminated many marine invertebrate groups and some fish lineages, but the largest arthrodires including Dunkleosteus survived into the Famennian.13 The final extinction of all placoderms came at the Devonian-Carboniferous boundary, approximately 359 million years ago, in an event that also devastated reef-building organisms and many other marine groups.13 The causes of the Late Devonian extinctions remain debated but likely involved a combination of oceanic anoxia, global cooling, and habitat loss associated with the spread of land plants and their effects on continental weathering and nutrient runoff.13 The disappearance of the placoderms opened ecological space that was subsequently filled by sharks and bony fishes, groups that have dominated aquatic ecosystems ever since.7, 13
References
Dunkleosteus terrelli (Placodermi: Arthrodira): a revision of the Cleveland Museum skull, with new data on cranial anatomy
Biomechanics of the jaw apparatus of the gigantic Devonian placoderm Dunkleosteus terrelli
Body size estimation in placoderms: comparison of methods and implications for the understanding of Devonian food webs