Overview
- The earliest tetrapods of the Late Devonian were polydactylous, semi-aquatic animals with functional gills and paddle-like limbs, but by the Late Carboniferous their descendants had radiated into dozens of lineages occupying freshwater, terrestrial, and even fully land-locked ecological niches.
- Romer's Gap, a 15-million-year interval of sparse tetrapod fossils bridging the Devonian and mid-Carboniferous, has been progressively filled by discoveries from Scotland and elsewhere, revealing that key terrestrial adaptations including pentadactyly and forward-facing feet arose during this poorly sampled period.
- By the end of the Carboniferous, tetrapods had diverged into temnospondyls, lepospondyls, and the amniote lineage, with early amniotes such as Hylonomus achieving full independence from water through the evolution of the cleidoic egg, setting the stage for the Permian radiation of reptiles and synapsids.
The emergence of limbed vertebrates from lobe-finned fishes during the Late Devonian period, roughly 380 to 360 million years ago, was only the beginning of the tetrapod story. The subsequent radiation of these early four-limbed animals through the Late Devonian, across the enigmatic interval known as Romer's Gap, and into the Carboniferous period produced a remarkable range of body plans and ecological strategies. From polydactylous, gill-bearing forms like Acanthostega that never left the water, to small insectivorous amniotes like Hylonomus that had severed all reproductive ties to aquatic habitats, the first 80 million years of tetrapod history established the major lineages that would dominate terrestrial ecosystems for the remainder of the Phanerozoic.1, 9
{kind=link}
Understanding early tetrapod diversity requires looking beyond the iconic fish-to-tetrapod transition itself and examining what happened after limbs with digits first appeared. The Devonian pioneers were just one chapter; the real diversification unfolded in the Carboniferous, when tetrapods exploited the vast new ecological space offered by the coal swamp forests that blanketed equatorial Pangaea.1, 10
Devonian pioneers
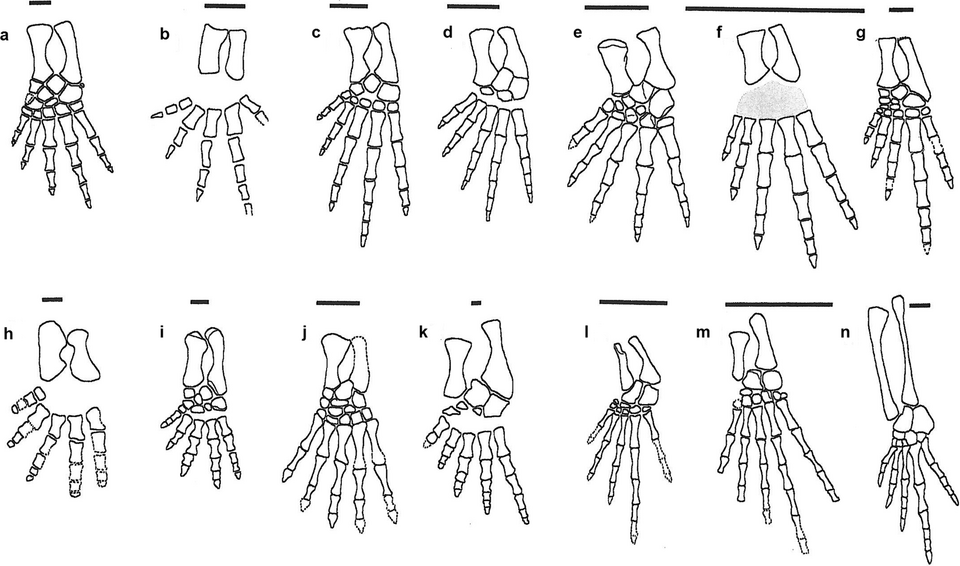
The oldest tetrapods known from body fossils date to the Famennian stage of the Late Devonian, approximately 365 to 360 million years ago. These animals were not a uniform group. Acanthostega gunnari, from East Greenland, possessed eight digits on each forelimb linked by webbing, retained a fish-like tail fin with lepidotrichia (bony fin rays), and bore well-ossified ceratobranchial bones with vascular grooves indicating functional internal gills for aquatic respiration.2, 3, 19 Its limbs lacked the robust wrist ossification needed for effective weight bearing on land, suggesting that digits initially served as paddles for navigating vegetated shallows rather than as instruments of terrestrial locomotion.3
Ichthyostega stensioei, roughly contemporary with Acanthostega and also from East Greenland, presented a strikingly different morphology despite its similar age and provenance. Reanalysis of its axial skeleton revealed broadly flanged, overlapping ribs that would have severely restricted lateral undulation of the trunk, and a regionalized vertebral column with a lumbar region adapted for dorsoventral flexion.4 With seven digits on its hind limb, Ichthyostega probably moved more like a seal than a salamander, using its powerful forelimbs to haul itself over substrates in a manner fundamentally different from the paddling locomotion of Acanthostega.1, 4
Tulerpeton curtum, from the Famennian of Tula in central Russia, added further variety. It possessed six digits on both fore and hind limbs and showed a more derived limb morphology than either of the Greenland genera, with features suggesting greater capacity for weight bearing.5 Phylogenetic analyses have placed Tulerpeton closer to the crown group of tetrapods than either Acanthostega or Ichthyostega, hinting that the divergence of major tetrapod lineages may have already been underway before the end of the Devonian.5, 9 Additional Devonian tetrapods known from fragmentary remains — including Ventastega from Latvia, Metaxygnathus from Australia, and Jakubsonia from Russia — confirm that by the latest Devonian, limbed vertebrates had achieved a near-global distribution across both Laurussia and Gondwana.1, 9
Romer's Gap and its resolution
Following the relatively diverse Devonian tetrapod assemblages, the fossil record becomes strikingly sparse for approximately 15 to 20 million years, spanning from the Devonian–Carboniferous boundary (359 Ma) through the Tournaisian stage of the Early Carboniferous (to roughly 345 Ma). This lacuna, identified by the American palaeontologist Alfred Sherwood Romer in the mid-twentieth century and known as Romer's Gap, long posed one of the most vexing problems in vertebrate palaeontology: when tetrapod fossils reappear in abundance in the mid-Visean, the fauna is utterly transformed. The polydactylous, aquatic Devonian forms have been replaced by animals with five or fewer digits, forward-facing feet, improved terrestrial locomotion, and recognisable affinities to the major lineages that would dominate the Carboniferous and beyond.1, 6
One influential explanation for the gap invokes atmospheric chemistry. Ward and colleagues marshalled geochemical evidence from sulfur isotope records and charcoal abundance to argue that atmospheric oxygen levels plunged to as low as 13 to 15 percent during the earliest Carboniferous, compared with approximately 21 percent today. Under such hypoxic conditions, air-breathing tetrapods dependent on inefficient buccal pumping would have been physiologically constrained, limiting their diversity and abundance and reducing their likelihood of entering the fossil record.7
The gap has, however, been progressively closed by new discoveries. In 2002, Jennifer Clack described Pederpes finneyae from the Tournaisian of Dumbarton, Scotland — the first articulated tetrapod skeleton from within Romer's Gap. Pederpes was functionally pentadactyl and possessed the earliest known forward-facing feet adapted for walking on land, bridging both the morphological and temporal divide between Devonian and mid-Carboniferous faunas.6 A decade later, Smithson and colleagues reported diverse tetrapod and arthropod assemblages from multiple Tournaisian localities in Scotland, including five new tetrapod taxa, demonstrating that the gap was at least partly a sampling artefact reflecting the rarity of suitable terrestrial sedimentary deposits rather than a genuine absence of tetrapod life.8 These Scottish faunas revealed that by the earliest Carboniferous, tetrapods had already begun to differentiate into distinct ecological guilds — aquatic, semi-aquatic, and terrestrial — with pelvic girdle morphology already showing the structural reorganisation necessary for weight-bearing locomotion on land.8, 12 These findings suggest that the critical innovations for land life were acquired rapidly in the wake of the Late Devonian extinction events.
Temnospondyls
The temnospondyls were the most species-rich and long-lived clade of early tetrapods, persisting from the Early Carboniferous to the Early Cretaceous — a span of more than 200 million years. They are first recognized in the fossil record from the Visean stage of the Carboniferous and rapidly diversified into a wide array of ecological types, from fully aquatic, flat-headed predators resembling modern crocodilians to small, salamander-like forms that inhabited moist forest floors.9, 10, 13
Carboniferous temnospondyls occupied a central role in the tetrapod faunas of equatorial coal swamp ecosystems. Genera such as Balanerpeton from the Visean of East Kirkton, Scotland, were relatively small, terrestrial animals with well-ossified limbs, while contemporaneous forms like Dendrerpeton from the Pennsylvanian of Joggins, Nova Scotia, are found inside fossilized tree stumps of the lycopsid Sigillaria, where they presumably sheltered or were trapped by flood waters.13, 20 Larger temnospondyls, including the Pennsylvanian genus Eryops, were robust, heavily built animals that functioned as apex predators in freshwater and marginal environments.10
The phylogenetic position of temnospondyls relative to modern amphibians remains one of the most debated questions in vertebrate systematics. One major hypothesis, supported by several phylogenetic analyses, holds that temnospondyls include the ancestors of modern lissamphibians — frogs, salamanders, and caecilians — making them the stem group from which all living amphibians descend.14 An alternative view places lissamphibians within or as close relatives of the lepospondyls, a second major group of early tetrapods. The resolution of this question has important implications for understanding how amphibian body plans evolved and when the key features of modern amphibians, such as metamorphosis and permeable skin, first appeared.9, 14
Lepospondyls
The lepospondyls were a diverse but generally small-bodied assemblage of Carboniferous and Permian tetrapods distinguished by their spool-shaped, directly ossified vertebral centra — in contrast to the multipartite vertebrae of temnospondyls. The group encompasses several distinct orders, including the snake-like aistopods, the aquatic nectrideans with their bizarre boomerang-shaped skulls, and the diminutive microsaurs, many of which were likely burrowing or leaf-litter-dwelling animals.10, 14
Whether lepospondyls constitute a natural (monophyletic) group or an artificial assemblage united only by shared primitive features has been a persistent question. Ruta and Coates's comprehensive phylogenetic analysis of early tetrapod relationships found support for lepospondyl monophyly in some analyses but not others, and the internal relationships of the group remain volatile depending on taxon sampling and character weighting.14 Regardless of their monophyly, lepospondyls played an important ecological role in Carboniferous and Early Permian ecosystems, occupying niches — burrowing, aquatic bottom-dwelling, elongate limbless locomotion — that temnospondyls generally did not exploit.10
The aistopods are among the most morphologically extreme early tetrapods. These elongate, entirely limbless animals, some exceeding a metre in length, are known from the Late Carboniferous and Early Permian. With up to 230 vertebrae and no trace of limbs or girdles in the most derived forms, aistopods converged on a body plan strikingly similar to that of modern snakes, representing one of the earliest experiments in limbless locomotion among tetrapods.10, 14
The origin and early diversification of amniotes
The most consequential event in Carboniferous tetrapod evolution was the origin of the amniotic egg, which freed vertebrates from the requirement to return to water for reproduction. The amniotic egg, with its protective membranes (amnion, chorion, allantois) and either a calcified shell or tough leathery covering, allowed embryos to develop in a self-contained aquatic environment on dry land. This innovation, which cannot be directly observed in the fossil record because eggs rarely fossilize, is inferred from the phylogenetic distribution of the amniotic condition among living vertebrates and from the skeletal features of the earliest fossil amniotes.10, 11
Casineria kiddi, from the Visean (approximately 340 Ma) of East Lothian, Scotland, is among the oldest fossils that may represent an amniote or a very close relative. Though known only from a partial postcranial skeleton, Casineria possesses gracile, well-ossified limbs with slender digits showing possible claws — features consistent with a fully terrestrial lifestyle and suggestive of amniote-grade terrestriality earlier than previously recognized.17 Its precise phylogenetic position remains uncertain, but its morphology indicates that the divergence between amniotes and non-amniote tetrapods was already underway by the mid-Carboniferous.1, 17
Hylonomus lyelli, from the late Pennsylvanian (approximately 312 Ma) of Joggins, Nova Scotia, is the oldest animal widely accepted as a true reptile and amniote. First described by John William Dawson in 1860, Hylonomus was a small, lizard-like animal approximately 20 centimetres in total length, with sharp conical teeth suited to an insectivorous diet. Its fossils were found inside the hollow stumps of lycopsid trees at the famous Joggins Fossil Cliffs, a UNESCO World Heritage Site, where they had apparently become trapped or sought shelter.11, 20 Robert Carroll's detailed 1964 description of the skull and postcranium confirmed Hylonomus as an early member of Reptilia, possessing features such as a fully consolidated braincase and an astragalus bone in the ankle that distinguish it from non-amniote tetrapods.11
By the Late Carboniferous, amniotes had diverged into at least two major lineages: the synapsids, which would eventually give rise to mammals, and the reptiles (including the diapsids, from which dinosaurs, birds, and modern reptiles descend). This fundamental split in the amniote family tree is documented by fossils from the Pennsylvanian of North America and Europe, and phylogenetic analyses suggest the divergence occurred no later than approximately 312 to 318 million years ago.10, 18 The rapidity with which amniotes diversified once they achieved reproductive independence from water underscores the transformative ecological significance of the amniotic egg.16
Adaptations for terrestrial life
The transition from water to land imposed a suite of physiological and anatomical challenges that early tetrapods addressed through a combination of inherited pre-adaptations and novel evolutionary innovations. The Devonian pioneers had already inherited lungs from their sarcopterygian ancestors — lungfishes, the closest living relatives of tetrapods, retain functional lungs to this day — but the earliest tetrapods still relied heavily on gills and cutaneous (skin-based) gas exchange. Acanthostega's retention of functional internal gills demonstrates that obligate air breathing came well after the origin of limbs.19 The loss of the operculum (the bony gill cover) in forms more derived than Panderichthys eliminated the opercular pump, the primary mechanism for ventilating gills, forcing increasing reliance on buccal pumping of air into the lungs.1
Limb and girdle strengthening progressed incrementally. The pectoral girdle, originally attached to the skull in fishes, was freed to create a mobile neck in Tiktaalik and all subsequent tetrapods. Through the Carboniferous, the pelvic girdle became increasingly robust and firmly attached to the vertebral column via a sacral rib connection, providing the structural support necessary for hind-limb-driven terrestrial locomotion. The consolidation of the pentadactyl limb — replacing the variable six-to-eight digit pattern of Devonian forms — appears to have been linked to improved mechanical efficiency for weight bearing and propulsion on land.1, 6
Skull modifications reflected the shift from aquatic suction feeding to terrestrial prey capture. The broad, flat skulls of many Devonian and early Carboniferous tetrapods, optimized for generating suction currents to engulf prey underwater, gave way in more terrestrial lineages to narrower, deeper skulls with stronger jaw musculature suited to biting and holding prey in air. The stapes bone, derived from the hyomandibular of fishes, was repurposed from jaw support to sound transmission, forming the basis of the middle ear in terrestrial tetrapods.1, 10
Carboniferous ecosystems and tetrapod ecology
The ecological theatre in which early tetrapod diversification played out was the Carboniferous world — a planet radically different from the modern one. Equatorial regions of Pangaea were blanketed by vast coal swamp forests dominated by giant lycopsids (Lepidodendron, Sigillaria), tree ferns, calamites, and seed ferns, forming dense, humid, multi-layered canopies over extensive wetlands. These forests were the primary habitat of most early tetrapods, and the diversity of ecological niches within them — forest floor, tree hollows, freshwater pools, swamp margins — provided the selective landscape that drove tetrapod diversification.10, 18
Atmospheric oxygen levels rose dramatically during the Carboniferous, reaching an estimated 30 to 35 percent by the Late Pennsylvanian compared with 21 percent today. This oxygen-rich atmosphere had profound consequences for terrestrial life. It enabled the evolution of giant arthropods, including griffinfly insects of the genus Meganeura with wingspans approaching 70 centimetres and millipede-like Arthropleura exceeding two metres in length.15 For tetrapods, elevated oxygen would have improved the efficiency of buccal pumping and cutaneous respiration, potentially facilitating the evolution of larger body sizes and more active metabolisms in lineages that were still refining their respiratory systems.7, 15
Tetrapod communities in the Pennsylvanian coal swamps were structured around waterways and wetland margins. Large temnospondyl amphibians served as the dominant aquatic and semi-aquatic predators, occupying ecological roles analogous to modern crocodilians. Smaller temnospondyls and lepospondyls filled a range of niches as aquatic insectivores, burrowers, and terrestrial foragers. The amniotes, initially a minor component of these faunas, increasingly exploited drier, more upland habitats as the Carboniferous progressed, a pattern documented by Sahney, Benton, and Ferry's analysis of Permo-Carboniferous terrestrial vertebrate diversity.18 The ability of amniotes to reproduce away from standing water gave them access to habitats that remained closed to amphibians, and by the end of the Carboniferous, early synapsids and reptiles had begun the ecological expansion that would accelerate dramatically in the Permian.10, 18
Significance of early tetrapod radiation
The Devonian-to-Carboniferous radiation of tetrapods established the fundamental blueprint for terrestrial vertebrate life. Within 80 million years of the first appearance of limbs with digits, tetrapods had produced aquatic predators, burrowing limbless forms, small terrestrial insectivores, and the first animals capable of completing their entire life cycle on dry land. This initial burst of morphological and ecological experimentation set the stage for every subsequent chapter of land vertebrate evolution: the Permian dominance of synapsids, the Mesozoic age of dinosaurs, and ultimately the rise of mammals in the Cenozoic.1, 10
The early tetrapod record also provides a compelling case study in how major evolutionary radiations unfold. The Devonian pioneers were morphologically diverse but ecologically constrained, tied to aquatic and marginal habitats. The passage through Romer's Gap — whatever its environmental causes — was accompanied by the acquisition of key innovations (pentadactyl limbs, improved respiration, the amniotic egg) that unlocked new adaptive zones. The subsequent Carboniferous explosion of lineages was not a simple linear progression but a branching radiation into multiple distinct body plans, many of which (aistopods, nectrideans, many temnospondyl families) later went extinct without leaving modern descendants.9, 14 The survivors — the lineage leading to lissamphibians and the amniote clade — represent only a fraction of the morphological experimentation that characterized the first great age of tetrapod diversity.
References
The Devonian tetrapod Acanthostega gunnari Jarvik: postcranial anatomy, basal tetrapod interrelationships and patterns of skeletal evolution
Confirmation of Romer’s Gap as a low oxygen interval constraining the timing of initial arthropod and vertebrate terrestrialization
Earliest Carboniferous tetrapod and arthropod faunas from Scotland populate Romer’s Gap
A new stem tetrapod from the Early Carboniferous of Scotland and the ancestral condition of the tetrapod pelvic girdle
Temnospondyl amphibians from the Early Carboniferous of Scotland and the role of metabolic rates in the radiation of tetrapods
Atmospheric oxygen, giant Paleozoic insects and the evolution of aerial locomotor performance
The origin and early radiation of the therapsid mammal-like reptiles: a palaeobiological hypothesis