Overview
- Canalization is the property of developmental systems to produce a consistent phenotype despite genetic and environmental perturbations, a concept introduced by C. H. Waddington in 1942 through his metaphor of the epigenetic landscape in which development flows along buffered channels toward stable outcomes.
- Waddington's experiments on Drosophila demonstrated genetic assimilation — the process by which an environmentally induced phenotype becomes genetically fixed after selection — providing early evidence that developmental buffering mechanisms can themselves evolve and that hidden genetic variation can be released when canalization breaks down.
- Modern molecular studies have identified specific canalization mechanisms, most notably the Hsp90 chaperone protein, which buffers the phenotypic effects of cryptic genetic variation under normal conditions but releases that variation under stress, potentially fuelling rapid evolutionary change in novel environments.
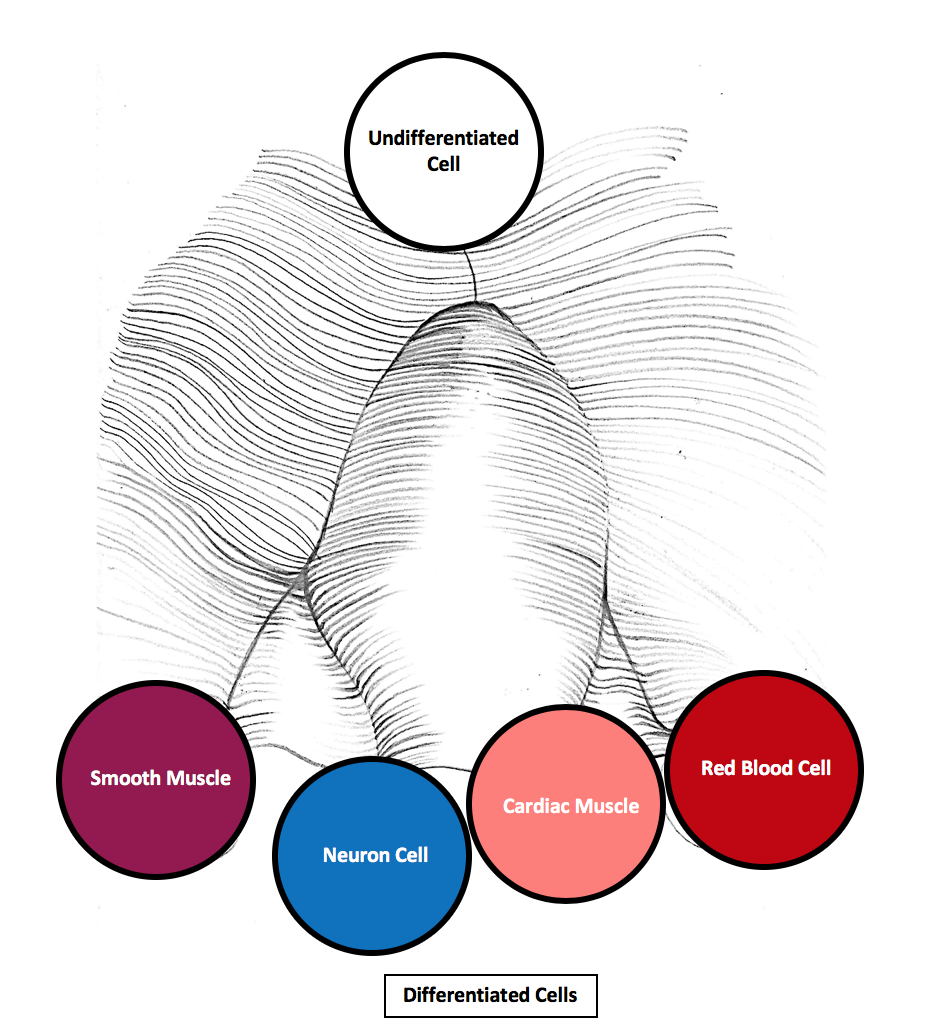
Canalization is the tendency of developmental systems to produce a consistent, invariant phenotype despite genetic mutations and environmental perturbations. The concept was introduced by the British developmental biologist C. H. Waddington in 1942, who proposed that natural selection favours developmental pathways that are buffered against disturbance, producing phenotypes that are robust to both environmental noise and the effects of new mutations.1 Waddington illustrated the idea with his famous metaphor of the epigenetic landscape: a surface of hills and valleys down which a ball (representing a developing cell or tissue) rolls toward one of several stable endpoints. The valleys represent canalised developmental trajectories, and the ridges between them represent the buffering mechanisms that prevent perturbations from pushing development into an alternative channel.3 Canalization is closely related to the modern concept of phenotypic robustness and has become central to discussions of evolvability, cryptic genetic variation, and the relationship between development and evolution.6, 13
{kind=link}
Waddington's experiments
Waddington's most influential experiments involved subjecting Drosophila melanogaster pupae to environmental shocks — heat shock or ether vapour — and selecting for individuals that displayed a particular phenotypic response. In his crossveinless experiment, he exposed pupae to a heat shock of 40°C for four hours during a critical developmental window and found that a small fraction of the resulting flies lacked the posterior crossvein on their wings. By selecting these crossveinless individuals as parents for the next generation and repeating the heat shock each generation, Waddington observed that the proportion of crossveinless flies increased over successive generations. After approximately 14 generations of selection, some flies produced the crossveinless phenotype even without the environmental stimulus.2
Waddington termed this process genetic assimilation: the conversion of an environmentally induced phenotype into a genetically determined one through selection. He interpreted the result as evidence that the wild-type crossvein phenotype is normally canalised — buffered by multiple genetic factors that ensure the crossvein develops correctly despite environmental variation. The heat shock overwhelmed this buffering, revealing underlying genetic variation that was normally hidden. Selection then shifted the genetic composition of the population so that the crossveinless phenotype could be produced without environmental perturbation.2, 3 Genetic assimilation does not invoke Lamarckian inheritance; the mechanism is entirely Darwinian, relying on selection acting on pre-existing genetic variation that is exposed when canalization is disrupted.11, 14
Molecular mechanisms of canalization
The molecular basis of canalization remained largely speculative until a landmark 1998 study by Rutherford and Lindquist demonstrated that the molecular chaperone Hsp90 acts as a capacitor of morphological evolution in Drosophila. Hsp90 is a heat-shock protein that assists in the correct folding of a wide range of signalling proteins, including many involved in developmental pathways. Under normal conditions, Hsp90 buffers the phenotypic effects of mutations in its client proteins by ensuring they fold and function correctly despite destabilising amino acid changes. When Hsp90 function is compromised — either by pharmacological inhibition with geldanamycin or by mutation in the Hsp83 gene — previously hidden genetic variation is expressed as a broad range of morphological abnormalities affecting eyes, legs, wings, and other structures.4
Queitsch, Sangster, and Lindquist extended these findings to Arabidopsis thaliana, showing that reduced Hsp90 function similarly reveals cryptic genetic variation affecting stem, leaf, and root morphology in plants, demonstrating that Hsp90-dependent canalization is not limited to animals.5 Importantly, some of the phenotypic variants revealed by Hsp90 reduction could be selected and stabilised in subsequent generations even when Hsp90 function was restored, paralleling Waddington's genetic assimilation result at a molecular level. These experiments provided the first concrete molecular mechanism for a canalisation system and suggested that stress-induced breakdown of chaperone-mediated buffering could release bursts of phenotypic variation upon which selection could act.4, 5
Beyond Hsp90, other canalization mechanisms have been identified. Redundant gene regulatory networks, in which multiple transcription factors regulate the same target gene, provide robustness because the loss of one regulator can be compensated by others. Von Dassow and colleagues demonstrated that the segment polarity gene network in Drosophila is remarkably robust to parameter variation, producing the correct striped pattern of gene expression across a wide range of kinetic rate constants.8 Gene duplication provides another source of robustness: when a gene is duplicated, mutations in one copy may be buffered by the continued function of the other.9, 15
Canalization and cryptic genetic variation
A central implication of canalization is that populations accumulate cryptic genetic variation — heritable variation that has no phenotypic effect under normal conditions because it is masked by developmental buffering. This hidden variation is not visible to natural selection and can therefore accumulate by genetic drift to levels that would be impossible if it were phenotypically expressed and exposed to purifying selection.12 When canalization breaks down, whether due to environmental stress, genetic perturbation, or entry into a novel environment, the cryptic variation is released and produces a burst of phenotypic diversity that selection can act upon.4, 12
The relationship between canalization and cryptic variation creates a potential mechanism for rapid evolutionary change. Under stable conditions, canalization maintains phenotypic constancy while genetic variation silently accumulates. Under stressful or novel conditions, the breakdown of canalization releases this stored variation, producing a wide array of phenotypes. If some of these phenotypes are advantageous in the new environment, selection can rapidly shift the population toward a new phenotypic optimum. In this view, canalization does not simply constrain evolution but also facilitates it by storing genetic variation that can be deployed when circumstances change.6, 10, 12
Evolution of canalization
Whether canalization evolves as a direct target of natural selection or as a by-product of stabilising selection on the phenotype has been debated since Waddington's original proposals. Waddington argued that canalization is itself an evolved property: populations in which developmental pathways are buffered against perturbation have higher mean fitness because fewer individuals deviate from the optimal phenotype. Stabilising selection thus favours genetic variants that reduce the phenotypic effects of environmental noise and new mutations, progressively deepening the valleys in the epigenetic landscape.1, 3 Theoretical models have generally supported this view, showing that stabilising selection can indeed lead to the evolution of reduced phenotypic variance, particularly when the fitness landscape is concave around the optimum.7
However, the evolution of canalization is complicated by its relationship to evolvability. A highly canalised system is robust to perturbation but may also be resistant to adaptive evolution because mutations have reduced phenotypic effects. This creates a potential tension between short-term robustness and long-term adaptability. Some theorists have proposed that fluctuating environments may favour intermediate levels of canalization that balance robustness against current perturbations with the capacity to respond to future environmental shifts.7, 10 Félix and Barkoulas have argued that phenotypic robustness and evolvability are not necessarily in conflict, because the cryptic genetic variation accumulated under canalization provides a reservoir of potential adaptive responses that can be released precisely when environmental conditions change and new phenotypes are needed.13
The study of canalization has moved from Waddington's conceptual framework to a molecularly grounded discipline that connects developmental biology, genetics, and evolutionary theory. The discovery of specific molecular capacitors such as Hsp90, the characterisation of robust gene regulatory network architectures, and the growing appreciation of cryptic genetic variation as an evolutionary resource have collectively established canalization as a key mechanism linking development to evolution. Understanding how organisms buffer their phenotypes against perturbation, and how that buffering can break down to enable rapid adaptive change, remains a central challenge at the intersection of evo-devo and population genetics.6, 10, 13