Overview
- Human childbirth is uniquely difficult among primates because bipedalism reshaped the pelvis into a narrow, twisted birth canal through which a large-brained fetus must rotate during delivery, creating what has been called the 'obstetric dilemma.'
- The classic obstetric dilemma hypothesis — that pelvic width is constrained by the demands of efficient bipedal locomotion — has been challenged by Holly Dunsworth's EGG (Energetics of Gestation and Growth) hypothesis, which argues that the timing of birth is driven by maternal metabolic limits rather than pelvic constraints.
- The rotational birth mechanism, obligate assisted delivery, and high rates of cephalopelvic disproportion in modern humans are evolutionary consequences of the intersection of two powerful selection pressures: efficient bipedal locomotion and encephalization.
Human childbirth is among the most dangerous and painful reproductive events in the animal kingdom. Unlike the relatively swift and uncomplicated deliveries typical of other primates, human birth involves a tight fit between the fetal head and the maternal pelvis, a complex series of rotations as the fetus navigates a tortuous birth canal, and a high risk of complications including obstructed labor, perineal tearing, and maternal or infant death.4, 10 These difficulties are not random misfortune. They are the predictable consequences of two of the most defining features of human evolution: bipedal locomotion, which reshaped the pelvis into a narrow, asymmetric passage, and encephalization, which expanded the fetal skull to dimensions that strain the limits of that passage.1, 7 Understanding the evolution of human childbirth requires grappling with how these two selection pressures have interacted over millions of years.
{kind=link}
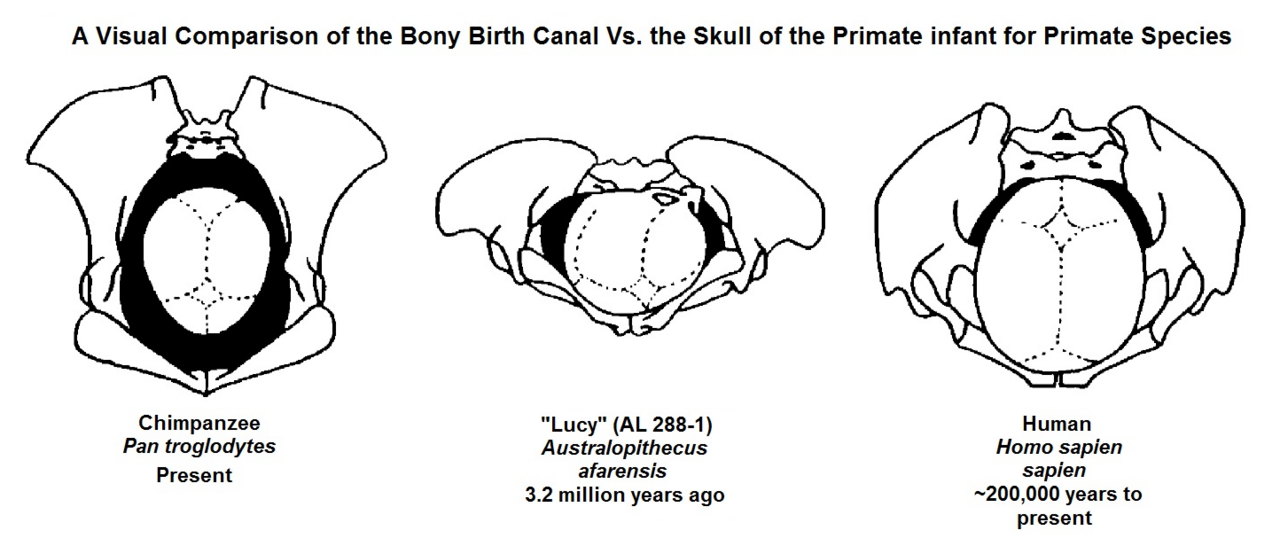
The obstetric dilemma
The concept of the "obstetric dilemma" was first articulated by Sherwood Washburn in 1960 and has dominated thinking about human birth for more than half a century. Washburn argued that bipedalism imposed two conflicting demands on the hominin pelvis. Efficient upright walking requires a narrow pelvis with short ilia (hip blades) and a compact pelvic outlet, which keeps the center of gravity close to the midline and reduces the energetic cost of locomotion. Successful childbirth, by contrast, requires a wide pelvic canal to accommodate the passage of a large-brained infant. According to Washburn, natural selection struck a compromise: the pelvis evolved to be just wide enough for a fetus to pass through, but the fetus was born at an earlier stage of brain development than would be ideal, resulting in the extreme helplessness (altriciality) of human newborns relative to other primate neonates.1
The obstetric dilemma framework makes several specific predictions. It predicts that there should be measurable locomotor costs associated with wider pelvises, that the human pelvis represents a biomechanical compromise between walking efficiency and birth canal size, and that the timing of human birth is constrained by the upper limit of fetal head size that can pass through the pelvic canal.1, 8 This framework became the standard textbook explanation for the difficulty of human childbirth and the apparent immaturity of human newborns. However, beginning in the 2000s, several of these predictions came under serious empirical challenge.2, 9
Pelvic morphology and the birth canal
The human pelvis differs from that of other great apes in ways that directly affect the birth process. In chimpanzees and gorillas, the birth canal is spacious relative to the neonatal head, and the inlet (the top of the pelvic opening) and outlet (the bottom) are both widest in the same dimension — the transverse plane. The fetal head enters and exits the canal in the same orientation, and delivery is typically rapid and uncomplicated.4, 8
In humans, the birth canal is both narrower and more complex. The pelvic inlet is widest in the transverse dimension (side to side), but the midpelvis is roughly circular, and the outlet is widest in the sagittal dimension (front to back). This means that the widest diameter of the birth canal rotates by approximately ninety degrees from inlet to outlet.5, 8 The human fetal head, which is longest in the sagittal dimension, must therefore enter the pelvis facing sideways (with the sagittal suture aligned transversely), rotate as it descends through the midpelvis, and emerge facing backward (with the sagittal suture aligned sagittally). This rotational birth mechanism is unique to humans among living primates.4, 8
The reorganization of the pelvis for bipedal locomotion fundamentally created this challenge. In quadrupedal primates, the ilia are long and flat, and the sacrum is positioned differently relative to the hip joints. The transition to habitual bipedalism shortened and broadened the ilia, repositioned the sacrum, and brought the ischial spines (bony projections on the inner pelvic wall) closer together, creating the narrow midpelvic plane that forces fetal rotation.5, 11 Reconstructions of the pelvis of Australopithecus afarensis suggest that even early hominins with relatively small brains (approximately 400 cubic centimeters) may have experienced some degree of tight fit between the fetal head and the birth canal, though likely without the full rotational mechanism seen in modern humans.6, 13
The EGG hypothesis
In 2012, Holly Dunsworth and colleagues published a direct challenge to the locomotor component of the obstetric dilemma. Their paper, titled "Metabolic hypothesis for human altriciality," argued that the timing of human birth is not determined by pelvic constraints on fetal head size but rather by maternal metabolic limits on gestation duration.2
Dunsworth's team, which called their framework the EGG hypothesis (Energetics of Gestation and Growth), presented several lines of evidence. First, they showed that the metabolic cost of human pregnancy follows a predictable trajectory that approaches the ceiling of sustained maternal metabolic output — approximately 2.1 times the basal metabolic rate — at roughly forty weeks of gestation. This metabolic ceiling, rather than pelvic constraints, appears to determine when birth occurs.2 Second, they demonstrated that human gestation length, when scaled for body mass, is not unusually short relative to other primates. Humans actually gestate longer than predicted for a primate of our size. What makes human neonates appear altricial is not premature birth but the enormous amount of postnatal brain growth — the human brain triples in size after birth, a pattern of postnatal growth that dwarfs that of other primates.2, 12
Third, a separate study by Anna Warrener and colleagues in 2015 tested the biomechanical prediction of the obstetric dilemma — that wider hips should impose locomotor costs — and found no such effect. In a series of experiments measuring the energetic costs of walking and running in individuals with varying pelvic breadth, wider hips did not result in measurably less efficient locomotion, undermining the premise that natural selection against wide pelvises has constrained birth canal size.9, 15
The EGG hypothesis does not deny that the pelvis constrains fetal head size at the moment of birth — the tight fit is an anatomical reality — but it reframes the question of why birth occurs when it does. Under the classic obstetric dilemma, gestation ends because the fetal head cannot get any larger and still fit through the pelvis. Under the EGG hypothesis, gestation ends because the mother can no longer sustain the metabolic cost of growing a fetus, and the tight fit at delivery is a consequence rather than the cause of birth timing.2, 7
Rotational birth and assisted delivery
Regardless of why birth occurs when it does, the mechanism of human birth is extraordinary among primates. In most primates, the neonate emerges facing toward the mother (occiput posterior presentation), allowing the mother to reach down, clear the infant's airway, and guide it toward the breast. In humans, the fetal head typically enters the pelvis in a transverse orientation and rotates during descent so that the infant emerges facing away from the mother (occiput anterior presentation). This backward-facing emergence makes it difficult for the mother to assist her own delivery without risking injury to the infant's spine or neck.4, 8, 10
Wenda Trevathan has argued that this rotational mechanism is the evolutionary basis for the near-universal human practice of assisted delivery — the presence of a birth attendant (midwife, partner, or relative) during labor. In other primates, mothers typically give birth alone. In humans, unassisted birth carries substantially elevated risk, and cross-cultural ethnographic evidence indicates that attended birth is the norm in virtually every human society.10 Rosenberg and Trevathan have proposed that obligate cooperative birthing is a derived human trait that evolved in concert with the increasing tightness of fit between fetal head and maternal pelvis over the course of Homo evolution, and that it represents one of the earliest forms of cooperative behavior in the hominin lineage.4
Evidence from Neanderthal pelvic anatomy suggests that rotational birth is not unique to Homo sapiens within the genus Homo. The Kebara 2 Neanderthal pelvis, one of the most complete archaic Homo pelvises known, shows a birth canal shape that would have required fetal rotation, though the specific pattern of rotation may have differed from that of modern humans.14 This implies that the basic rotational birth mechanism evolved before the divergence of the Neanderthal and modern human lineages, perhaps in Homo heidelbergensis or an earlier common ancestor, in response to increasing fetal brain size in the Middle Pleistocene.14
Cephalopelvic disproportion
The tight fit between fetal head and maternal pelvis in humans means that relatively small deviations from average dimensions — a slightly larger fetal head, a slightly narrower pelvis — can result in obstructed labor, a condition known as cephalopelvic disproportion (CPD). Before the advent of modern obstetric intervention, CPD was a leading cause of maternal and neonatal mortality. Even today, in regions with limited access to emergency cesarean section, obstructed labor remains a significant cause of maternal death and birth injury.17
Philipp Mitteroecker and colleagues have modeled the evolutionary dynamics of CPD and suggested that the widespread availability of cesarean delivery since the mid-twentieth century may have begun to shift selection pressures on pelvic and fetal head dimensions. Because cesarean section allows survival in cases of CPD that would previously have been fatal to mother and infant, it effectively relaxes the selection pressure that maintained the close match between fetal head and pelvic dimensions. Their model predicts a slow increase in the rate of CPD over evolutionary time — on the order of 10 to 20 percent over several generations — though the effect would be modest given the short time frame and the many other factors influencing pelvic and cranial dimensions.17
The rate of obstructed labor varies among human populations, reflecting both genetic variation in pelvic dimensions and environmental factors such as maternal nutrition, adolescent growth patterns, and fetal macrosomia (excessive fetal size, often associated with gestational diabetes). Populations with shorter average stature and smaller pelvic dimensions experience higher rates of CPD, as do populations where nutritional improvement has increased average birth weight faster than pelvic dimensions have responded.7
Evolutionary trajectory
The difficulty of human childbirth did not arise all at once. It emerged incrementally over millions of years as different hominin lineages evolved increasing degrees of bipedal efficiency and brain size. The earliest bipeds, such as Australopithecus afarensis, had small brains (approximately 400 to 500 cubic centimeters) but already possessed a pelvis significantly reshaped for upright walking. Reconstructions suggest a moderately tight fit between the neonatal head and the birth canal, but probably without the full rotational mechanism of modern humans.6, 13
The transition to larger brains in early Homo (approximately 600 to 900 cubic centimeters) would have further tightened the relationship between fetal head and pelvis, likely requiring some degree of fetal rotation. By the time of Homo heidelbergensis and early Homo sapiens, with brain sizes approaching 1,200 to 1,500 cubic centimeters, the full modern pattern of rotational birth was probably established.5, 14 The fossil evidence is sparse — complete pelvises are rare in the hominin record — but what exists is consistent with a gradual tightening of the obstetric fit across Homo evolution.5
Modern humans are thus the product of an evolutionary history in which two of the most powerful selection pressures — for efficient locomotion and for large brains — pulled pelvic morphology in opposing directions. The result is not a graceful solution but an imperfect compromise: a birth canal that is just barely adequate, a fetus that must execute a complex rotational maneuver to pass through it, and a neonate so neurologically immature that it requires years of intensive care before it can function independently.7, 18 The evolution of cooperative breeding, extended parental investment, and cultural transmission of obstetric knowledge can all be understood as downstream consequences of this fundamental anatomical constraint.4, 10
References
The evolution of the human pelvis: changing adaptations to bipedalism, obstetrics and thermoregulation
The evolution of the human pelvis: its evolutionary relationships and their implications for the onset of bipedalism