Overview
- Ammonites were a spectacularly diverse subclass of cephalopod molluscs (Ammonoidea) that originated in the Early Devonian around 409 million years ago and persisted for roughly 340 million years until the end-Cretaceous mass extinction 66 million years ago, producing more than 10,000 described species across eight or more orders.
- Their chambered shells, divided internally by septa whose junction with the outer wall formed increasingly complex suture patterns — from simple goniatitic lines in the Paleozoic to intricately frilled ammonitic sutures in the Mesozoic — provide one of the most detailed records of morphological evolution in the fossil record.
- Because ammonite species evolved rapidly and were distributed globally in marine environments, they serve as the premier index fossils for subdividing and correlating Mesozoic rocks, with more than 1,500 biostratigraphic zones defined on the basis of ammonite assemblages across the Jurassic and Cretaceous alone.
Ammonites — more precisely ammonoids, members of the subclass Ammonoidea — were an extraordinarily diverse group of cephalopod molluscs that dominated the world's oceans for over 340 million years, from their origin in the Early Devonian approximately 409 million years ago to their final extinction at the Cretaceous-Paleogene boundary 66 million years ago.4, 12 Characterized by their coiled, chambered shells, ammonoids were close relatives of modern coleoid cephalopods (squid, octopuses, and cuttlefish) rather than of the externally similar nautiluses, and they produced more than 10,000 described species — and likely many thousands more — distributed across virtually every marine environment on every continent.1, 22 Their rapid evolutionary turnover, global distribution, and distinctive shell morphology make them among the most important index fossils in the geological record, indispensable for dating and correlating Mesozoic marine strata worldwide.11
Shell morphology and the chambered conch
The ammonoid shell, or conch, is a hollow tube coiled into a planispiral (flat spiral) in most species, though some groups evolved radically aberrant shapes including straight, helical, U-shaped, and paperclip-like forms.17, 8 The shell is divided internally into a series of gas-filled chambers (camerae) separated by thin walls called septa. The animal occupied only the outermost chamber — the body chamber — while the inner chambers functioned as a buoyancy apparatus.9 A fleshy tube called the siphuncle threaded through all the septa, connecting the chambers and enabling the animal to regulate the ratio of gas to liquid within them, thereby controlling its buoyancy in the water column.3, 9

{kind=link}
The external surface of the conch displays a suite of morphological features that are fundamental to classification. The umbilicus is the depression at the centre of coiling; shells range from involute (where later whorls overlap earlier ones, producing a narrow umbilicus) to evolute (where whorls barely overlap, leaving a wide, open umbilicus).17 The venter is the outer margin of the whorl, which may be rounded, flattened, or keeled. The aperture is the opening of the body chamber from which the animal's soft parts emerged; in some species the aperture bore lateral extensions called lappets or a ventral projection called a rostrum.17, 1 Shell ornamentation — including ribs, tubercles, spines, and growth lines — varies enormously across taxa and provides primary characters for species identification.1
Shell size ranged from species smaller than one centimetre in diameter to the enormous Parapuzosia seppenradensis from the Late Cretaceous, which reached diameters exceeding 1.7 metres and is among the largest shelled animals ever to have lived.1, 22
Suture patterns and their evolution
The line where each septum meets the inner surface of the outer shell wall is called a suture, and suture patterns are among the most taxonomically and evolutionarily informative features of ammonoid shells. Because sutures are visible on internal moulds (steinkerns) — the most common mode of ammonoid preservation — they have been central to ammonoid classification since the earliest days of palaeontology.6, 7 Suture patterns became progressively more complex through ammonoid evolutionary history, a trend that represents one of the clearest examples of increasing morphological complexity in the fossil record.6
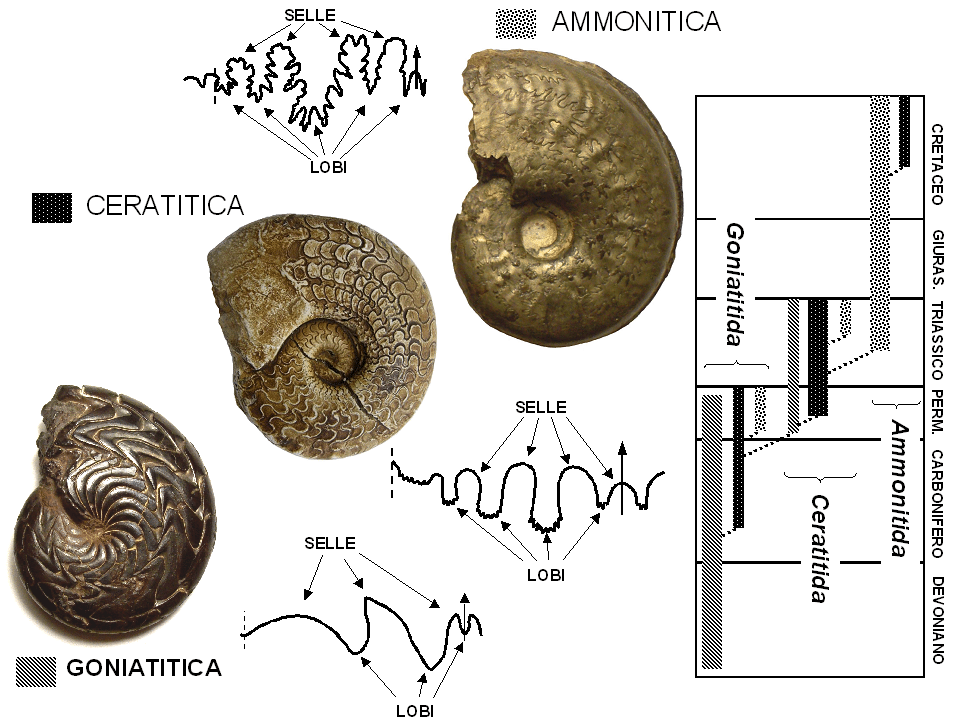
{kind=link}
Three fundamental suture types are recognized. Goniatitic sutures, characteristic of Paleozoic ammonoids (order Goniatitida and relatives), consist of smooth, undivided lobes (downward-pointing folds) and saddles (upward-pointing folds), typically numbering fewer than eight elements.6, 1 Ceratitic sutures, typical of many Triassic ammonoids (order Ceratitida), feature serrated lobes but smooth, rounded saddles, representing an intermediate level of complexity.1, 6 Ammonitic sutures, characteristic of Jurassic and Cretaceous ammonites (orders Ammonitida, Phylloceratida, and Lytoceratida), display both lobes and saddles that are intricately subdivided into secondary and tertiary elements, producing fractal-like patterns of extraordinary complexity.6, 7
The functional significance of increasingly complex sutures remains debated. Hypotheses include strengthening the shell against hydrostatic pressure at depth, improving buoyancy regulation by increasing the surface area available for cameral liquid exchange, and enhancing resistance to predatory attacks. Inoue and Kondo proposed that complex suture patterns may be a by-product of reaction-diffusion mechanisms in the rear mantle tissue rather than a directly selected adaptation, suggesting that morphogenetic processes rather than functional demands may have driven suture elaboration.7
Soft-body anatomy and mode of life
Because ammonoid soft tissues are preserved only in the rarest of circumstances, reconstructing their anatomy and ecology depends heavily on inference from shell morphology, comparison with living nautiluses and coleoid cephalopods, and the handful of exceptional fossils that preserve soft parts.3, 14 The animal occupied the body chamber with its head and tentacles protruding from the aperture. Recent studies of exceptionally preserved specimens from the Late Jurassic Solnhofen Limestone of Germany have revealed details of the brachial crown (arm apparatus), showing that ammonoids bore arm-like structures equipped with hook-like armatures, differing substantially from the smooth, numerous tentacles of the living nautilus.14
A remarkable perisphinctid specimen from the early Tithonian of Eichstätt, Germany, preserved in isolation from its shell, revealed a complete digestive tract, paired gills within a mantle cavity, a hyponome (jet-propulsion funnel), and what appear to be male reproductive organs — providing the most complete picture of ammonoid internal anatomy available from the fossil record.15 The presence of a hyponome confirms that ammonoids, like living cephalopods, used jet propulsion for locomotion, expelling water from the mantle cavity through the funnel to generate thrust.10, 15
Buoyancy calculations based on three-dimensional shell reconstructions indicate that most normally coiled ammonoids achieved near-neutral buoyancy, enabling them to float in the water column with minimal expenditure of energy.9 The orientation of the living animal within the water column depended on the distribution of mass within the shell: species with longer body chambers tended to hang with the aperture pointing downward, while those with shorter body chambers assumed a more horizontal posture with the aperture directed forward, facilitating active swimming.9, 10 Ammonoids are generally considered to have been slower and less manoeuvrable than coleoid cephalopods, which had internalized or lost their shells entirely, but faster and more agile than the living nautilus, whose relatively simple siphuncle limits its buoyancy regulation.10
Sexual dimorphism
Many ammonoid species exhibit pronounced sexual dimorphism, expressed as two distinct adult morphotypes occurring in the same stratigraphic beds and sharing identical juvenile growth trajectories before diverging at maturity.16 The larger form, termed the macroconch, is interpreted as the female and typically possesses a simple, unmodified aperture. The smaller form, the microconch, is interpreted as the male and commonly develops lateral projections called lappets or ventral extensions at the aperture, features that may have played a role in mating.16, 3
Size ratios between macroconchs and microconchs vary across taxa but are frequently in the range of 2:1 to 3:1 in diameter, with the female body chamber providing substantially more internal volume — presumably for egg production.16 The recognition that pairs of formerly separate "species" actually represent sexual dimorphs of a single biological species has significantly revised ammonoid taxonomy, reducing the number of valid species in many lineages. De Baets and colleagues documented cases where macroconch-microconch assignments have been reversed upon closer examination, cautioning that dimorphic pairing must be supported by shared ontogenetic trajectories and stratigraphic co-occurrence rather than assumed from size differences alone.16
Heteromorph ammonoids
While the majority of ammonoid species bore tightly coiled planispiral shells, a significant minority evolved radically different shell geometries collectively termed heteromorphs. Heteromorph ammonoids abandoned the conventional spiral in favour of straight (orthoconic), helically coiled (turreted), U-shaped (ancyloconic), hook-shaped, or irregularly uncoiled shells.8 Heteromorphic coiling evolved independently in multiple ammonoid lineages across their entire evolutionary history, but reached its greatest diversity during the Cretaceous, when the order Ancyloceratina produced a spectacular radiation of unconventionally shaped forms including Baculites (straight), Turrilites (helical), Nipponites (irregularly tangled), and Scaphites (partially uncoiled).8, 12

_(Cretaceous).jpg){kind=link}
Hoffmann and colleagues demonstrated that heteromorph ammonoids could achieve near-neutral buoyancy regardless of their shell shape, contradicting earlier assumptions that aberrant coiling represented degeneracy or pathological forms on the brink of extinction.8, 9 Straight-shelled forms like Baculites likely assumed a vertical orientation in the water column with the aperture pointing downward, functioning as ambush predators or plankton feeders drifting in midwater.8 Helically coiled forms occupied ecological niches with no close analogue among living cephalopods. Far from being evolutionary dead ends, heteromorphs were ecologically successful and in some Cretaceous formations constitute the majority of ammonoid specimens.8, 12
Evolutionary history
Ammonoids originated in the Early Devonian (Emsian stage, approximately 409 million years ago) from straight-shelled cephalopods belonging to the order Bactritida. The evolutionary transition from bactritid to ammonoid is documented by a series of transitional forms from the Tafilalt region of Morocco, showing a progressive increase in shell curvature from the straight Bactrites through the loosely curved Lobobactrites and Cyrtobactrites to the loosely coiled Anetoceras, the earliest true ammonoid.4, 21 The coiling of the shell into a spiral improved hydrodynamic stability and enabled more efficient buoyancy regulation, advantages that may have driven the rapid diversification of early ammonoids.5
During the remainder of the Paleozoic, ammonoids diversified through three major radiations punctuated by severe extinction events. The goniatites dominated from the Devonian through the Permian, producing forms with simple goniatitic sutures that radiated extensively in Carboniferous tropical seas.22, 1 The end-Permian mass extinction approximately 252 million years ago — the most severe biotic crisis in Earth's history — devastated ammonoid diversity, reducing the subclass to a handful of surviving lineages.19 The recovery was remarkably rapid: within the first one to two million years of the Triassic, ammonoids had reradiated into a diverse array of ceratites bearing ceratitic suture patterns, recolonizing marine habitats worldwide at a pace that ranks among the fastest known faunal recoveries in the fossil record.19
The end-Triassic extinction again reduced ammonoid diversity, but the survivors gave rise to the great Jurassic and Cretaceous radiation of true ammonites — forms characterized by highly complex ammonitic sutures and extraordinary morphological diversity.2, 22 Ammonites reached their peak diversity during the middle to late Cretaceous, when they occupied habitats ranging from shallow continental shelves to deep oceanic waters, from tropical to polar seas.12, 22
Ammonoid diversity through geological time22
Biostratigraphic importance
Ammonoids are widely regarded as the single most important group of index fossils for the Mesozoic Era. An effective index fossil must be morphologically distinctive, geographically widespread, temporally short-ranged, and abundantly preserved — criteria that ammonoids satisfy to an exceptional degree.11 Individual ammonoid species typically persisted for time intervals of 0.5 to 2 million years before evolving into morphologically distinct successors, a turnover rate fast enough to subdivide geological time into fine-grained biostratigraphic zones while being slow enough that each species had time to spread across wide geographic areas.11, 2
The Jurassic and Cretaceous periods are subdivided into a dense network of ammonite biozones — more than 1,500 in total — that provide temporal resolution finer than any other biostratigraphic scheme available for Mesozoic marine rocks.11 Each biozone is defined by the presence of one or more characteristic ammonite species, and the sequential replacement of these zones records the pace of evolutionary change through time. Because many ammonite species were distributed across multiple oceanic basins, their biozones enable correlation of strata between continents that were widely separated during the Mesozoic, linking, for example, Jurassic sections in England with coeval deposits in East Africa, South America, and the Himalayas.11, 2
In the Paleozoic, goniatites and related ammonoids performed a similar biostratigraphic role, particularly for the Devonian and Carboniferous, where ammonoid-based zonation supplements or replaces conodont and brachiopod schemes in many regions.22, 1 The biostratigraphic utility of ammonoids stems ultimately from the same biological properties that made them ecologically successful: their rapid reproduction, planktonic larval dispersal, and evolutionary responsiveness to environmental change ensured that new species arose frequently and spread quickly.2
Extinction at the Cretaceous-Paleogene boundary
Ammonoids vanished entirely during the end-Cretaceous mass extinction 66 million years ago, an event triggered by the impact of a roughly 10-kilometre asteroid at Chicxulub on the Yucatán Peninsula, which produced catastrophic global environmental disruption including impact winter, wildfires, acid rain, and the collapse of marine food webs.18, 12 At least 57 ammonoid species belonging to six superfamilies were alive during the final 500,000 years of the Cretaceous, demonstrating that the group remained diverse and ecologically significant up to the very end — contrary to earlier narratives that portrayed ammonoids as already declining before the impact.12
The selectivity of the extinction has long puzzled researchers: ammonoids were annihilated, yet the superficially similar nautiloids survived. Tajika and colleagues proposed that the critical difference lay in metabolism and reproductive strategy. Ammonoids are inferred to have had higher metabolic rates and produced numerous small, planktonic hatchlings that drifted in surface waters — the very habitat most devastated by the post-impact collapse of primary productivity.13 Nautiloids, by contrast, produced fewer, larger eggs that hatched as benthic juveniles, bypassing the lethal surface-water conditions. The combination of high metabolic demands (requiring continuous food intake) and vulnerable planktonic larvae may have rendered ammonoids fatally susceptible to the brief but intense disruption of ocean productivity that followed the Chicxulub impact.13
Evidence from Antarctica and other high-latitude sites suggests that the extinction may have proceeded in two phases: an initial decline possibly linked to Deccan Traps volcanism in the final hundreds of thousands of years of the Cretaceous, followed by the coup de grâce delivered by the bolide impact.20 Regardless of whether the extinction was abrupt or phased, no ammonoid is reliably documented from rocks younger than the earliest Danian, and the subclass Ammonoidea — one of the most successful and long-lived groups of marine animals in Earth's history — ended its 340-million-year reign at the K-Pg boundary.12, 18
Scientific and cultural legacy
Ammonoids occupy a central place in the history of palaeontology and geology. They were among the first fossils to be recognized as the remains of once-living organisms, and their orderly succession in sedimentary strata was instrumental in the development of the principle of faunal succession — the foundational insight that different rock layers contain different assemblages of fossils, enabling the relative dating of geological formations.11, 2 William Smith's pioneering geological map of England and Wales (1815), which demonstrated that strata could be identified and correlated by their fossil content, relied heavily on ammonite distributions, establishing a method that remains central to stratigraphy today.11
The name "ammonite" derives from the Egyptian god Ammon (Amun), whose ram-like coiled horns the fossils were thought to resemble. In medieval Europe, ammonites were known as "snakestones" and attributed magical properties — a cultural association commemorated in the coat of arms of Whitby, Yorkshire, where Jurassic ammonites are abundant in the coastal cliffs.3 Today, ammonoids remain among the most intensively studied fossil groups, with their shells providing data on ancient ocean temperatures (via oxygen isotope analysis), water depth, ocean circulation patterns, and the tempo of evolutionary change. The two comprehensive volumes of Ammonoid Paleobiology edited by Klug and colleagues in 2015 synthesize the current state of knowledge, covering anatomy, ecology, diversity, macroevolution, and paleogeography across the full 340-million-year span of ammonoid history.1, 2
References
Understanding ammonoid sutures: new insight into the dynamic evolution of Paleozoic suture morphology
Ammonites on the brink of extinction: diversity, abundance, and ecology of the order Ammonoidea at the Cretaceous/Paleogene (K/Pg) boundary
New evidence from exceptionally 'well-preserved' specimens sheds light on the structure of the ammonite brachial crown
Failed prey or peculiar necrolysis? Isolated ammonite soft body from the Late Jurassic of Eichstätt (Germany) with complete digestive tract and male reproductive organs
Another lesson from beautiful monsters: the case of 'sex reversals' in the Ammonoidea and their significance
The Chicxulub asteroid impact and mass extinction at the Cretaceous-Paleogene boundary
Nautiloids before and during the origin of ammonoids in a Siluro-Devonian section in the Tafilalt, Anti-Atlas, Morocco