Overview
- Azhdarchidae were a family of Late Cretaceous pterosaurs that included the largest flying animals ever to exist, with genera such as Quetzalcoatlus and Hatzegopteryx reaching wingspans of 10–12 metres and standing heights rivalling modern giraffes.
- Anatomical evidence—including elongated limbs, generalist skulls, hyper-elongate cervical vertebrae with a unique spoke-like internal architecture, and a strong continental fossil bias—supports the hypothesis that azhdarchids were terrestrial stalkers that foraged on foot like giant storks or ground hornbills.
- On the Late Cretaceous island of Hațeg (modern Romania), the robust, short-necked azhdarchid Hatzegopteryx occupied the ecological role of apex predator in the absence of large theropod dinosaurs, demonstrating the remarkable ecological versatility of the group before its extinction at the Cretaceous–Paleogene boundary 66 million years ago.
Azhdarchidae are a family of pterodactyloid pterosaurs that dominated Late Cretaceous skies from approximately 90 to 66 million years ago. Named after Azhdarcho lancicollis, a species described by Lev Nesov in 1984 from the Bissekty Formation of Uzbekistan, the family takes its name from azhdar, a dragon-like creature in Persian mythology.6 Azhdarchids include the largest flying animals known to science: genera such as Quetzalcoatlus, Hatzegopteryx, and Arambourgiania achieved wingspans of 10 to 12 metres and, when standing on the ground, would have looked a giraffe in the eye.7, 16 Yet the family also encompassed smaller members with wingspans as modest as 2 to 3 metres, revealing a breadth of ecological diversity that extended far beyond sheer size.2
The azhdarchid body plan was distinctive among pterosaurs. All members shared proportionally large, edentulous (toothless) skulls atop hyper-elongate cervical vertebrae, relatively short wings with an abbreviated fourth phalanx, and unusually long hindlimbs suited to quadrupedal terrestrial locomotion.1, 16 This combination of features set azhdarchids apart from most other pterosaur lineages and generated decades of debate over how these animals fed and lived. In recent years, an accumulating body of evidence has converged on a striking conclusion: azhdarchids were predominantly terrestrial foragers, striding across floodplains and savannahs much like oversized storks or ground hornbills, supplementing this lifestyle with efficient soaring flight across vast distances.1, 10
Discovery and naming
The first azhdarchid material to attract scientific attention was a single enormous cervical vertebra collected in 1943 from the Maastrichtian phosphates of Jordan and described in 1954 as Titanopteryx philadelphiae (later renamed Arambourgiania philadelphiae).2, 16 For decades, however, the significance of this specimen was not fully appreciated. The formal recognition of Azhdarchidae as a distinct clade came only in 1984, when Nesov erected the subfamily Azhdarchinae (initially placed within Pteranodontidae) on the basis of over 200 fragmentary bones of Azhdarcho lancicollis from the Late Cretaceous of Uzbekistan, noting the animal's extremely elongated, cylindrical cervical vertebrae as a defining character.6
The discovery that truly thrust azhdarchids into public consciousness was Douglas Lawson's 1975 description of Quetzalcoatlus northropi from the Maastrichtian Javelina Formation of Big Bend National Park, Texas. Named for the Aztec feathered serpent deity Quetzalcoatl and the aviation pioneer John Northrop, this giant was represented by a partial wing comprising fragments of an enormously elongated fourth metacarpal and wing phalanx.5, 13 Subsequent collecting expeditions in the 1970s and 1980s recovered dozens of specimens of a smaller species, later designated Quetzalcoatlus lawsoni, whose more complete skeletal remains provided the foundation for understanding azhdarchid anatomy in detail.5, 13 In 2021, a landmark series of six papers published as a memoir of the Society of Vertebrate Paleontology finally described these materials comprehensively, nearly half a century after their discovery.12, 13
Other significant azhdarchid discoveries have expanded the family's geographic and temporal range considerably. Hatzegopteryx thambema was described in 2002 from the Maastrichtian Densu-Ciula Formation of Romania's Hațeg Basin, distinguished by an exceptionally wide skull, robust cervical vertebrae, and a spongy internal bone texture unlike the thin-walled hollow bones typical of most pterosaurs.15, 17 Alanqa saharica was named from the Cenomanian Kem Kem beds of Morocco, representing one of the earliest known azhdarchids.11 Ongoing fieldwork continues to reveal new species: azhdarchid remains have been reported from the Bayanshiree Formation of Mongolia, the Turonian of Utah, and even the terminal Cretaceous of Syria.14, 19, 20
Anatomy and skeletal structure
The azhdarchid skeleton is built around a set of shared anatomical characters that distinguish the family from other pterosaur groups. The skull was proportionally large relative to body size, elongate, and completely toothless, with a tapering, lance-shaped rostrum and jaws that could open to a wide gape.1, 16 In most azhdarchids the skull bore a low sagittal crest, and the nasoantorbital fenestra (a large opening combining the nasal and antorbital openings) occupied a substantial portion of the skull's lateral surface, reducing weight while maintaining structural rigidity.16

{kind=link}
The cervical vertebrae are the most diagnostic feature of the group. Azhdarchid neck vertebrae are extraordinarily elongated — individual centra can be more than eight times longer than they are wide — and are cylindrical in cross section rather than being dorsoventrally flattened as in many other pterodactyloids.1, 6 A 2021 study using X-ray computed tomography on Alanqa cervical vertebrae revealed a remarkable internal micro-architecture: the neural canal is incorporated within the body of the vertebra, creating a tube-within-a-tube arrangement, with the inner and outer tubes connected by helically arranged, spoke-like trabeculae resembling the spokes of a bicycle wheel.4 Finite element analysis demonstrated that as few as 50 of these trabeculae increased the buckling resistance of the vertebra by up to 90 percent, allowing the neck to bear substantial compressive and bending loads despite being extremely lightweight.4 This architecture explains how azhdarchids could support necks up to 3 metres long on bodies weighing only 200 to 250 kilograms.4, 7
The postcranial skeleton reflected a dual commitment to both flight and terrestrial locomotion. The wings were relatively short and broad compared to those of other large pterosaurs such as pteranodontids, with a calculated aspect ratio of approximately 8.1 — comparable to that of storks, vultures, and other large soaring birds that rely on thermal updrafts and slope soaring rather than dynamic soaring over open ocean.7 The fourth wing phalanx was markedly abbreviated, contributing to the short, broad wing planform.1 The hindlimbs were proportionally long for a pterosaur, and the overall limb proportions resembled those of modern large-bodied terrestrial birds more than those of other pterosaur families, suggesting substantial walking ability.1
Diversity and global distribution
Azhdarchids achieved a near-global distribution during the Late Cretaceous. Fossils have been recovered from every continent except Australia and Antarctica (where the absence may reflect sampling gaps rather than true absence), spanning a temporal range from the Turonian (approximately 93 million years ago) to the end-Maastrichtian (66 million years ago).2 They were the most diverse and widespread pterosaur family during the final 30 million years of the Cretaceous, and among the last pterosaurs to survive before the end-Cretaceous mass extinction extinguished the entire order.2, 16
A comprehensive 2014 review catalogued 54 azhdarchid occurrences worldwide, drawing from both skeletal remains and ichnofossils (trackways).2 The depositional settings of these occurrences reveal a broad environmental tolerance: 35 percent derive from coastal marine sediments, 18 percent from estuarine and lagoonal settings, 17 percent from coastal plains, 17 percent from fluvial plains, and 13 percent from lacustrine deposits.2 This environmental breadth stands in contrast to the strong continental bias of azhdarchid fossils — they are disproportionately found in terrestrial and marginal-marine facies rather than deep-marine settings, a pattern consistent with a primarily terrestrial lifestyle supplemented by coastal and inland foraging.1, 2
Depositional settings of azhdarchid fossil occurrences2
Major azhdarchid-bearing localities include the Javelina and Aguja formations of Texas, the Kem Kem beds of Morocco, the Hațeg Basin of Romania, the Bissekty Formation of Uzbekistan, the Dinosaur Park Formation of Alberta, the Bayanshiree Formation of Mongolia, and phosphatic deposits of Jordan and Syria.2, 9, 11, 14, 19, 20 Recent discoveries continue to extend the known range: azhdarchid material from the Turonian of Utah represents one of the oldest North American occurrences, while 2025 reports from Syria and Mongolia have added new data on Late Cretaceous diversity in regions previously undersampled for pterosaurs.14, 19, 20
The terrestrial stalking hypothesis
For much of the twentieth century, azhdarchids were assumed to have been fish-eaters that plucked prey from water surfaces while in flight, analogous to modern skimmers or pelicans.1 This interpretation began to unravel with a 2008 study by Mark Witton and Darren Naish, who systematically evaluated azhdarchid functional morphology and concluded that the evidence overwhelmingly favoured a terrestrial foraging ecology.1 Their "terrestrial stalking" hypothesis proposed that azhdarchids were predominantly ground-based predators and foragers that walked quadrupedally across open and partially wooded landscapes, using their long necks and beaks to seize small animals, invertebrates, and possibly carrion from the ground — a lifestyle most closely analogous to that of modern ground hornbills and large storks such as the marabou.1
Multiple independent lines of evidence support this hypothesis. First, the arthrological range of motion in azhdarchid cervical vertebrae indicates that the neck was best suited for dorsoventral (up-and-down) movements rather than the lateral sweeping motions that would be required for skim-feeding or surface-dipping.1, 10 Second, azhdarchid skulls lack the specialised jaw morphologies found in known piscivorous pterosaurs: there are no tooth-like projections, no laterally compressed mandibles, and no kinetic joints that would facilitate rapid jaw closure on aquatic prey.1 Third, the proportionally long hindlimbs and compact feet of azhdarchids indicate a narrow, efficient quadrupedal gait well suited to sustained walking, and pterosaur trackways attributed to azhdarchids confirm terrestrial proficiency.1, 10 Fourth, the strong continental skew in azhdarchid fossil occurrences — with the majority found in terrestrial and marginal-marine sediments rather than open-marine settings — is consistent with animals that spent most of their time on land.1, 2
A 2015 follow-up study by Witton and Naish addressed subsequent criticisms, including an alternative "pelican-mimic" hypothesis that proposed azhdarchids waded in shallow water and scooped fish with flexible throat pouches. They found no anatomical basis for a pelican-like throat pouch in azhdarchid skulls and reaffirmed that the totality of evidence — skeletal proportions, cranial mechanics, depositional context, and ichnological data — most parsimoniously supported terrestrial stalking as the primary foraging strategy.10
Flight and launch biomechanics
Despite their enormous size, azhdarchids were capable fliers. Biomechanical analyses indicate that the largest species, with wingspans approaching 10 to 12 metres and estimated body masses of 200 to 250 kilograms, operated near but not beyond the theoretical limits of powered flight in a terrestrial vertebrate.7, 18 Their wing aspect ratio of approximately 8.1, combined with relatively high wing loading, indicates that azhdarchids were specialist soarers rather than continuous flappers, exploiting thermals, ridge lift, and other atmospheric phenomena to stay aloft for extended periods with minimal energetic cost — much like modern vultures and albatrosses.7
The question of how such massive animals became airborne was resolved by Michael Habib's quadrupedal launch hypothesis. Habib (2008) observed that in all living tetrapod fliers, the number of limbs used for terrestrial locomotion matches the number used for launch: bipedal birds launch with their hindlimbs, while quadrupedal bats launch with all four limbs.8 Because pterosaurs were obligate quadrupeds on the ground, Habib proposed that they launched using a powerful vault from all four limbs, with the massively developed forelimbs — which bore the flight musculature — providing the primary thrust. Analysis of humeral and femoral bone strength in pterosaurs confirmed that the forelimbs were far more robust than the hindlimbs, consistent with a forelimb-dominated launch.8 For a 250-kilogram azhdarchid, this mechanism would have produced an initial leap of approximately 2.5 metres into the air, sufficient to clear the ground and begin the first wingbeat without the need for a running start, a cliff edge, or favourable headwinds.8, 12
The 2021 Quetzalcoatlus monograph refined these estimates further. Padian and colleagues modelled the flight envelope of Q. northropi and concluded that it could sustain soaring flight at speeds of approximately 90 kilometres per hour, with a cruising altitude that took advantage of high-altitude thermals available over the warm Cretaceous continental interiors.12 Once airborne, a giant azhdarchid could have covered hundreds of kilometres in a single flight, rivalling the long-distance soaring performance of modern condors and albatrosses.7, 12
Hatzegopteryx and the Hațeg Island ecosystem
Perhaps the most remarkable chapter in azhdarchid paleoecology concerns Hatzegopteryx thambema, a giant azhdarchid from the latest Cretaceous of Romania's Hațeg Basin.15, 17 During the Maastrichtian, the Hațeg region was an island within the Tethys archipelago — a chain of islands formed by tectonic uplift across what is now southern Europe. The island's fauna exhibited pronounced insular dwarfism: sauropod dinosaurs such as Magyarosaurus were reduced to the size of cattle, and hadrosaurids like Telmatosaurus were similarly diminutive.3, 9 Crucially, large theropod dinosaurs — the typical apex predators of Cretaceous terrestrial ecosystems — were absent or extremely rare on the island.3
In this predator-depleted environment, Hatzegopteryx appears to have filled the apex predator niche. A 2017 study by Naish and Witton examined the cervical vertebrae of Hatzegopteryx and found that, unlike the hyper-elongate necks of other azhdarchids, Hatzegopteryx possessed a proportionally short, massively robust neck approximately 1.5 metres long, with cervical vertebrae bearing extensive muscle attachment sites and a dense, spongy internal bone texture rather than the thin cortical walls typical of pterosaurs.3
{kind=link}
The skull was enormously wide — approximately half a metre across the temporal region — and equipped with powerful jaw-closing musculature.15, 17 Together, these features indicate an animal capable of handling large prey items, potentially including the dwarf dinosaurs that shared its island habitat.3
The Hațeg ecosystem thus provides a natural experiment in ecological role reversal: in the absence of theropod competition, a flying reptile adapted the azhdarchid body plan into that of a robust, ground-based macro-predator. This scenario is without modern parallel but underscores the ecological versatility that made azhdarchids the most successful pterosaur lineage of the final Cretaceous.3, 9
Key genera and size range
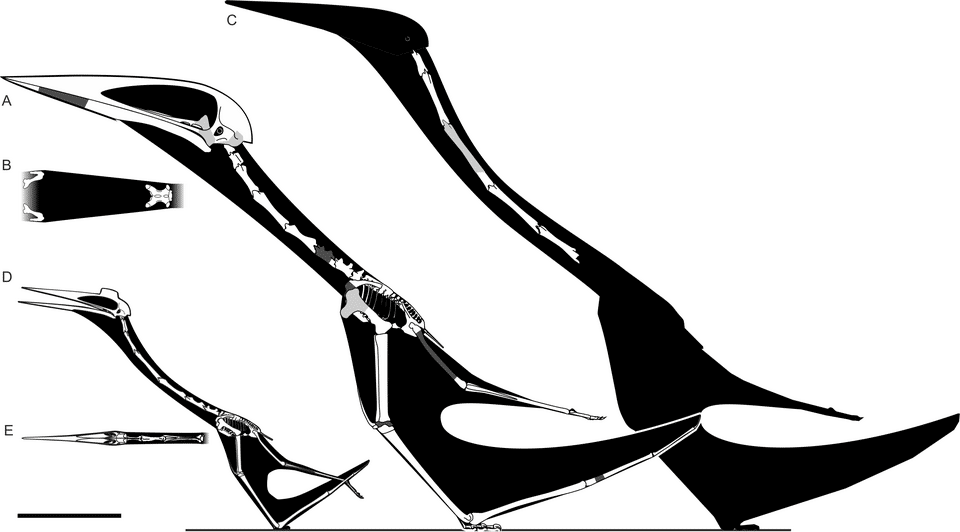
The family Azhdarchidae encompassed a wide range of body sizes. The largest known members — Quetzalcoatlus northropi, Hatzegopteryx thambema, and Arambourgiania philadelphiae — reached estimated wingspans of 10 to 12 metres and standing heights of approximately 5 metres, making them by far the largest flying animals in Earth's history.7, 13, 16 At the other end of the spectrum, the smaller species Quetzalcoatlus lawsoni had a wingspan of approximately 4.5 to 5.5 metres, and the early azhdarchid Azhdarcho lancicollis is estimated at roughly 3 to 4 metres wingspan.5, 6 Even smaller azhdarchids have been reported from the Cretaceous of Hungary and Romania, with estimated wingspans as low as 2 to 3 metres.2, 9

_(Solnhofen_Limestone,_Upper_Jurassic;_Bavaria,_Germany)_(36543746860).jpg){kind=link}
Selected azhdarchid genera and estimated wingspans2, 5, 7, 11, 13, 16
| Genus | Age | Region | Est. wingspan |
|---|---|---|---|
| Quetzalcoatlus northropi | Maastrichtian | Texas, USA | 10–11 m |
| Hatzegopteryx thambema | Maastrichtian | Romania | 10–12 m |
| Arambourgiania philadelphiae | Maastrichtian | Jordan | ~10 m |
| Quetzalcoatlus lawsoni | Maastrichtian | Texas, USA | 4.5–5.5 m |
| Alanqa saharica | Cenomanian | Morocco | ~6 m |
| Azhdarcho lancicollis | Turonian | Uzbekistan | 3–4 m |
Evidence from the Javelina Formation of Texas suggests that multiple azhdarchid species of different body sizes coexisted within the same ecosystem, with variation in jaw morphology implying dietary niche partitioning.5, 13 Similar size-structured azhdarchid communities have been documented in the Hațeg Basin and in the Late Cretaceous of Morocco and Central Asia, suggesting that ecological partitioning by body size was a recurrent pattern in the family.2, 9, 11
Phylogenetic relationships
Azhdarchidae belong to the superfamily Azhdarchoidea, a diverse clade of derived pterodactyloid pterosaurs that also includes the families Tapejaridae, Thalassodromidae, and Chaoyangopteridae.5, 16 Within Azhdarchoidea, chaoyangopterids are generally recovered as the closest relatives of azhdarchids, sharing elongated cervical vertebrae and reduced dentition, though chaoyangopterids are known primarily from the Early Cretaceous of China and lack the extreme neck elongation of true azhdarchids.16
Internal phylogenetic relationships within Azhdarchidae remain the subject of active research. The 2021 phylogenetic analysis by Andres and Langston recovered two principal subclades: Quetzalcoatlini, comprising taxa more closely related to Quetzalcoatlus, and Hatzegopterygia, comprising taxa closer to Hatzegopteryx.5 Under this framework, the robust, short-necked morphology of Hatzegopteryx represents a derived specialisation within one subclade rather than the ancestral condition for the family as a whole.3, 5 Several genera once assigned to Azhdarchidae, including Alanqa and Argentinadraco, have been recovered in some analyses as members of related but distinct families, highlighting the ongoing instability in azhdarchoid systematics.2, 5
The broader phylogenetic position of pterosaurs within Archosauria has also seen recent revision. A 2020 study by Ezcurra and colleagues identified a series of Triassic archosaur precursors (lagerpetids) as the closest known relatives of Pterosauria, helping to bridge the morphological gap between terrestrial archosaurs and the earliest flying pterosaurs, though a substantial ghost lineage persists between these forms and the first appearance of azhdarchids in the Late Cretaceous.21
Extinction
Azhdarchids were among the last pterosaurs to inhabit Earth. By the late Maastrichtian, they were the only pterosaur family with a confirmed global presence, as other lineages such as pteranodontids and nyctosaurids had already declined or disappeared.2, 16 The end-Cretaceous mass extinction, triggered by the Chicxulub asteroid impact 66 million years ago, extinguished the entire order Pterosauria along with the non-avian dinosaurs, mosasaurs, ammonites, and many other groups.2
Whether azhdarchid diversity was already in decline before the impact event remains debated. Some palaeontologists have pointed to the apparent reduction in pterosaur species richness through the Late Cretaceous as evidence for a long-term decline driven by competition with increasingly diverse birds. Others have argued that this pattern is an artefact of the poor and geographically biased pterosaur fossil record, noting that azhdarchids remained ecologically diverse and geographically widespread right up to the Cretaceous–Paleogene boundary.7, 16 The recent description of new azhdarchid material from the latest Cretaceous of Syria and Mongolia supports the latter interpretation, suggesting that azhdarchids were still radiating into new regions and maintaining ecological diversity in the final millions of years before the impact.19, 20
Regardless of whether azhdarchid diversity was waning, the Chicxulub impact and its aftermath — global darkness, temperature collapse, the shutdown of photosynthesis, and cascading ecosystem failure — proved fatal. As large-bodied terrestrial foragers dependent on productive continental ecosystems, azhdarchids would have been especially vulnerable to the abrupt destruction of food webs that followed the impact.2, 16 With their extinction, 165 million years of pterosaur evolution came to an end, and the skies were left to birds alone.
References
Review of taxonomy, geographic distribution, and paleoenvironments of Azhdarchidae (Pterosauria)
Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators
Helically arranged cross struts in azhdarchid pterosaur cervical vertebrae and their biomechanical implications
Phylogenetic systematics of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea)
The osteology of Azhdarcho lancicollis Nessov, 1984 (Pterosauria, Azhdarchidae) from the Late Cretaceous of Uzbekistan
On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness
A New Azhdarchid Pterosaur from the Late Cretaceous of the Transylvanian Basin, Romania: Implications for Azhdarchid Diversity and Distribution
New azhdarchid pterosaur (Pterosauria, Azhdarchidae) from the Upper Cretaceous of Morocco
The anatomy and taxonomy of Quetzalcoatlus Lawson 1975 (Pterodactyloidea: Azhdarchoidea)
Hatzegopteryx sp. (Pterosauria, Azhdarchidae) from the Late Cretaceous of the Hațeg Basin, Romania
Recovering lost time in Syria: a gigantic latest Cretaceous azhdarchid pterosaur from the Palmyrides mountain chain
Azhdarchid pterosaur diversity in the Bayanshiree Formation, Upper Cretaceous of the Gobi Desert, Mongolia