Overview
- The argument from poor design (dysteleological argument) contends that numerous biological structures — the recurrent laryngeal nerve, the human vertebral column, the retinal blind spot, the pharynx serving both respiration and swallowing — exhibit features that would not be expected from an intelligent designer but are precisely what natural selection acting on inherited variation would produce.
- These structures are not merely suboptimal but bear the unmistakable signature of historical contingency: they make sense as modifications of ancestral forms under evolutionary constraints, but are inexplicable as the products of unconstrained purposeful engineering by an omnipotent, omniscient creator.
- Theistic responses include the appeal to unknown purposes (we cannot know why God designed things this way), the fall (design was originally perfect but corrupted by sin), and the claim that optimality is not a requirement of design — critics argue that each response either makes God’s design indistinguishable from no design at all or introduces unfalsifiable ad hoc assumptions.
The argument from poor design — also known as the dysteleological argument — contends that numerous features of the biological world exhibit structural deficiencies, inefficiencies, and hazards that are inconsistent with the hypothesis of an omniscient, omnipotent designer but are precisely what one would expect from the undirected process of evolution by natural selection. The argument does not claim that biological organisms are non-functional; they are, on the whole, remarkably effective at surviving and reproducing. The claim is that they bear the unmistakable marks of historical contingency, developmental constraints, and evolutionary tinkering — characteristics of a process that modifies existing structures rather than designing from scratch, and that optimizes for reproductive fitness rather than for engineering elegance.1, 2, 5
The argument has a long history in the philosophy of biology, predating Darwin. David Hume noted in Dialogues Concerning Natural Religion (1779) that the world contains enough suffering and apparent malfunction to cast doubt on the inference from order to a benevolent designer. After Darwin provided a mechanism — natural selection — that explains the appearance of design without invoking a designer, the argument from poor design became a specific, empirically grounded challenge to teleological arguments for God’s existence. It has been developed by Stephen Jay Gould, Richard Dawkins, Elliott Sober, Neil Shubin, Nathan Lents, and others, and it targets both classical natural theology and the modern intelligent design movement.2, 6, 11
The structure of the argument
The argument from poor design is typically formulated as an inference to the best explanation. It does not claim to prove that no designer exists; it claims that the specific features of biological organisms are better explained by evolution than by intelligent design. The logic runs as follows:5, 11
P1. If biological organisms were designed by an omniscient, omnipotent creator, we would expect them to exhibit efficient, elegant, and purpose-fitted design without unnecessary structural compromises.
P2. Many biological organisms exhibit structures that are inefficient, hazardous, or functionally compromised in ways that reflect historical constraints rather than purposeful engineering.
P3. These structures are precisely what we would expect from evolution by natural selection, which modifies inherited structures incrementally and cannot start from scratch.
C. Therefore, the features of biological organisms are better explained by evolution than by intelligent design.
The argument’s strength lies in the specificity of its predictions. Evolution by natural selection makes a distinctive prediction: organisms should bear the marks of their evolutionary history. Structures that evolved for one function and were later co-opted for another should retain traces of their original form. Developmental pathways inherited from ancestors should constrain the range of possible designs, even when the ancestral constraint is no longer functional. Vestigial structures — remnants of features that were useful in ancestral species but serve no current function — should be common. In each case, the evolutionary prediction is confirmed by the evidence, and the design hypothesis must resort to ad hoc explanations to accommodate the same evidence.1, 7, 14
Examples from anatomy
The most frequently cited example of poor design is the recurrent laryngeal nerve, particularly in the giraffe. This nerve connects the brain to the larynx, a distance of only a few inches in most mammals. However, instead of taking the direct route, the nerve descends from the brain into the thorax, loops around the aortic arch (in left-sided mammals) or the subclavian artery, and then ascends back up to the larynx. In the giraffe, this detour adds approximately 4.6 meters (15 feet) of unnecessary nerve length. The nerve’s circuitous path is a direct consequence of its evolutionary origin: in fish, the nerve serves the sixth branchial arch and passes behind the aortic arch in a direct route. As the mammalian neck elongated over evolutionary time, the nerve was progressively stretched around the arch because natural selection could not “rewire” the nerve to take a more direct path without first disconnecting it entirely — a lethal modification.3, 4, 14
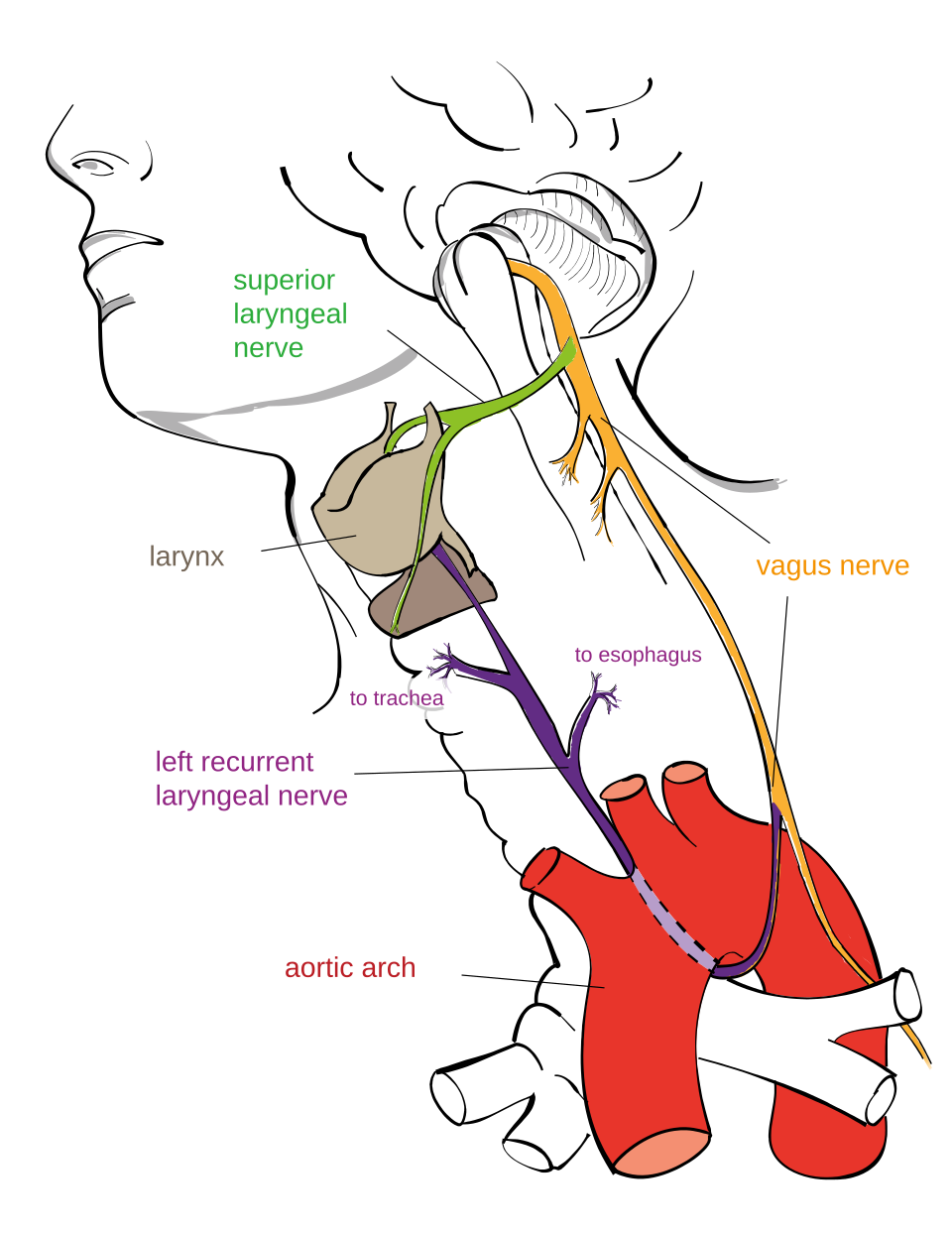
{kind=link}
The human vertebral column provides another striking example. The human spine is a modified version of the horizontal spine found in quadrupedal ancestors, repurposed for upright bipedal locomotion. This evolutionary history accounts for the high prevalence of lower back pain, herniated discs, and spinal stenosis in humans: the lumbar region bears compressive loads it was not “designed” for but was incrementally adapted to support. An engineer designing a bipedal organism from scratch would not produce a curved, segmented column with vulnerable intervertebral discs; the design reflects modification of an inherited quadrupedal plan rather than purposeful engineering for upright posture.14, 17
The vertebrate eye, often cited by design advocates as an example of irreducible complexity, actually illustrates poor design when examined closely. The mammalian retina is “inverted” — the photoreceptor cells face away from incoming light, with the nerve fibers and blood vessels layered on top, creating a blind spot where the optic nerve exits the retina. Cephalopod mollusks (octopuses and squid) evolved camera eyes independently, and their retinas are “right-side out,” with photoreceptors facing the light and no blind spot. The vertebrate arrangement is a consequence of the embryological development of the retina from the neural tube, an inherited developmental pathway that could not be reversed by natural selection. An intelligent designer, unconstrained by developmental history, would have no reason to install the retina backwards.1, 2, 17
Additional examples abound. The human pharynx serves as a shared pathway for both respiration and swallowing, creating a choking hazard that kills thousands of people annually — an arrangement inherited from fish, where the shared pathway posed no risk because fish do not swallow food into their lungs. The human reproductive and urinary systems share structures in a way that increases the risk of infection, particularly in females. The vas deferens in human males loops over the ureter rather than taking the direct path to the urethra, paralleling the recurrent laryngeal nerve in its unnecessary detour. Wisdom teeth, the appendix, the coccyx, and goosebumps (the contraction of arrector pili muscles attached to vestigial body hair) are all remnants of ancestral features that serve little or no current function.14, 17
Examples from genetics
The genomic evidence for poor design is, if anything, more striking than the anatomical evidence. The human genome is approximately 3.2 billion base pairs long, but only about 1.5% codes for proteins. Much of the remaining sequence consists of transposable elements (mobile genetic parasites that copy themselves throughout the genome), pseudogenes (broken copies of formerly functional genes), and repetitive sequences with no known function. The human genome contains more than one million copies of Alu elements alone — short sequences derived from a retrotransposon that has been replicating within primate genomes for tens of millions of years. This genomic architecture is exactly what evolution predicts: a genome that has accumulated vast amounts of non-functional or parasitic DNA over deep time, constrained by the inability of natural selection to purge sequences that impose only a small fitness cost.7, 17
Pseudogenes are particularly significant for the argument from poor design. Humans carry the gene for L-gulonolactone oxidase (GULO), the enzyme required for vitamin C synthesis, but the gene is broken by accumulated mutations and does not produce a functional protein. This is why humans require dietary vitamin C and can develop scurvy. The same gene is broken in the same way in other primates, indicating a shared mutation inherited from a common ancestor. If humans were designed from scratch, there would be no reason to include a broken vitamin C gene — but evolution predicts exactly this pattern, because mutations that disable a gene are preserved if the organism obtains the relevant nutrient from its diet (as fruit-eating primates do) and natural selection does not remove the broken gene.17, 14
Shared genetic errors across species provide some of the strongest evidence for common descent and against independent design. Humans and other great apes share identical inactivating mutations in multiple olfactory receptor genes, vitamin synthesis genes, and retroviral insertion sites. The probability that an intelligent designer would independently insert identical “broken” genes and identical retroviral sequences into the genomes of separately created species, in exactly the positions predicted by the phylogenetic tree, is vanishingly small. The evolutionary explanation — that these species inherited the shared errors from a common ancestor — is both simpler and more predictive.1, 7
Theistic responses
Theistic defenders of design have offered several responses to the argument from poor design. The most common is the appeal to unknown purposes: what appears to be poor design may in fact serve a purpose that humans have not yet discovered. William Dembski has argued that the inference from apparent suboptimality to actual suboptimality is unreliable, because our understanding of biological systems is incomplete, and structures that seem non-functional may turn out to have functions not yet identified. This response is structurally identical to the skeptical theist response to the evidential problem of evil: just as the skeptical theist argues that God may have morally sufficient reasons for permitting suffering that we cannot grasp, the design advocate argues that God may have design purposes for apparently suboptimal structures that we cannot identify.8, 11
Critics argue that this response makes the design hypothesis unfalsifiable. If every apparent instance of poor design can be explained away by appeal to unknown purposes, then no observation could ever count against the design hypothesis, and the hypothesis loses its explanatory power. Elliott Sober has made this point formally: a hypothesis that is compatible with every possible observation predicts nothing, and a hypothesis that predicts nothing explains nothing. The evolutionary hypothesis, by contrast, makes specific, testable predictions about the kinds of suboptimal structures organisms should exhibit — predictions that are confirmed by the evidence. The fact that evolution predicts the recurrent laryngeal nerve’s path while design theory can only accommodate it after the fact is an evidential asymmetry that favors evolution.5, 16
A second theistic response appeals to the biblical Fall: God’s original creation was perfect, but the introduction of sin through human disobedience corrupted the natural world, introducing disease, suffering, and structural degradation. On this view, poor design is not a reflection of God’s creative intentions but a consequence of the curse described in Genesis 3. This response faces several difficulties. It requires a literal reading of the Genesis narrative that is inconsistent with the scientific evidence for the age of the Earth and the deep history of life. Many of the “design flaws” in organisms are shared with species that, on any theological account, predate the existence of humans — the recurrent laryngeal nerve is present in fish, which are not descendants of Adam. And the response cannot account for the systematic pattern of the flaws: they are not random degradations but precisely the kinds of historical constraints that evolutionary theory predicts.10, 13
A third response, advanced by some proponents of intelligent design, is that optimality is not a necessary feature of design. Human engineers produce suboptimal designs all the time — constrained by budgets, materials, deadlines, and competing requirements — and we do not conclude that their products are undesigned. Michael Behe has argued that the relevant question is not whether organisms are optimally designed but whether they exhibit “irreducible complexity” — systems composed of interacting parts that could not have been assembled by gradual addition. On this view, poor design is compatible with intelligent design as long as the overall system could not have arisen through undirected evolution.9
This response concedes more than its proponents may intend. Human engineers are constrained by material, temporal, and cognitive limitations; an omnipotent, omniscient designer has none of these constraints. If the designer is God as classically conceived — unlimited in power, knowledge, and time — there is no reason for suboptimal design. The analogy with human engineering works only if the designer is conceived as limited, which conflicts with the theological commitments of classical theism. Furthermore, Behe’s concept of irreducible complexity has been challenged on empirical grounds: systems that appear irreducibly complex, such as the bacterial flagellum, have been shown to contain components that function in other contexts, suggesting evolutionary pathways through co-option and modification of existing parts.10, 12, 13
Philosophical significance
The argument from poor design occupies a specific niche in the landscape of arguments against theism. It is not a general argument against the existence of God; it is a targeted argument against the inference from biological complexity to an intelligent designer. It leaves open the possibility that God exists but did not directly design biological organisms — a position held by theistic evolutionists such as Kenneth Miller, who accepts the full evolutionary account of biological diversity while maintaining that God created the universe and its laws. On this view, poor design is not an argument against God’s existence but against the specific claim that individual organisms or structures were purposefully engineered by a divine intelligence.12, 18
The argument also has implications for the fine-tuning argument. Proponents of fine-tuning argue that the physical constants of the universe are calibrated with extraordinary precision to permit life, suggesting a cosmic designer. The argument from poor design does not directly address cosmic fine-tuning, but it raises a methodological question: if the same designer who fine-tuned the constants of physics also designed biological organisms, why is the cosmic design apparently exquisite while the biological design is riddled with inefficiencies and hazards? The tension between cosmic fine-tuning and biological poor design suggests that either the designer had different standards for physics and biology (an ad hoc assumption) or that the fine-tuning of physical constants and the structure of biological organisms have different explanations.5, 11, 15
For the broader debate between theism and naturalism, the argument from poor design functions as part of a cumulative case. No single instance of suboptimal structure is decisive, but the pervasive pattern of evolutionary tinkering across the entire tree of life — from genomic parasites to anatomical detours to developmental constraints — constitutes a body of evidence that is naturally explained by evolution and that design theory must explain away case by case. The cumulative weight of this evidence, combined with the absence of any biological structure that unambiguously requires a designer, supports the conclusion that the appearance of design in biology is the product of natural selection rather than intelligent agency.1, 6, 18