Overview
- Conodonts are tiny, tooth-like phosphatic elements that are among the most abundant and biostratigraphically useful fossils in the Paleozoic and Triassic, spanning approximately 300 million years from the late Cambrian to the end-Triassic extinction (~495–201 Ma).
- The nature of the conodont animal remained enigmatic for over 150 years until the discovery of soft-bodied conodont specimens in the 1980s and 1990s, which revealed eel-like chordates roughly 40 mm long with large eyes, a notochord, V-shaped myomeres, and a feeding apparatus of conodont elements in the head region.
- Conodont elements underwent rapid evolutionary change and had wide geographic distributions, making them among the most powerful index fossils for Paleozoic biostratigraphy, conodont alteration indices also serve as geothermometers for assessing the thermal maturity of sedimentary rocks.
The conodont enigma
For more than 150 years following their first description by Christian Heinrich Pander in 1856, conodonts were among the most enigmatic fossils in paleontology. The elements themselves — tiny, tooth-like structures composed of calcium phosphate (apatite), typically 0.2 to 5 mm in length — were found in enormous numbers in marine sedimentary rocks spanning from the late Cambrian to the end of the Triassic, a stratigraphic range of approximately 300 million years.6, 13 Their biological affinities remained wholly unclear: different workers variously assigned them to annelid worms, fish, molluscs, arthropods, chaetognaths, and even plants. The elements were manifestly organic hard parts, but without knowledge of the animal that bore them, their systematic position was irresolvable.5, 7
{kind=link}
Discovery of the conodont animal
The breakthrough came in the 1980s and 1990s with the discovery of exceptionally preserved soft-bodied conodont animals. The first convincing specimen was recovered from the Lower Carboniferous (Visean) Granton Shrimp Bed near Edinburgh, Scotland, described by Briggs, Clarkson, and Aldridge in 1983.4, 10 This specimen preserved the outline of a small, elongate, eel-like animal approximately 40 mm long with a conodont apparatus in the head region, a pair of large eyes, a notochord, and V-shaped muscle blocks (myomeres) along the trunk — features that collectively identified the animal as a chordate.3, 4
Subsequent discoveries from the Silurian of Wisconsin and additional Carboniferous localities confirmed and refined this picture.1 The South African Soom Shale (Ordovician) yielded large conodont animals up to 40 cm in length, demonstrating greater size variation than the Scottish specimens suggested.11 All known conodont body fossils share the same basic bauplan: an elongate, laterally compressed body with a terminal or subterminal mouth, large paired eyes, a notochord running the length of the body, chevron-shaped myomeres, a caudal fin, and a bilaterally symmetric feeding apparatus of conodont elements in the pharyngeal region.2, 11
Phylogenetic position
The discovery of the conodont animal resolved the century-long debate about affinity but opened a new one: where exactly do conodonts fit within Chordata? The presence of a notochord, myomeres, and large paired eyes clearly places them among chordates, but whether they are vertebrates, stem vertebrates, or more basal chordates has been debated.2, 15
Donoghue and colleagues argued that conodont elements are homologous to vertebrate teeth and contain tissues (enamel and dentine) that are uniquely vertebrate characters, placing conodonts among crown or stem vertebrates.2, 6 Others have questioned whether the mineralized tissues in conodont elements are truly homologous to vertebrate enamel and dentine or represent convergent evolution of hard tissues in a more basal chordate lineage.9 The current consensus favors a position for conodonts as stem gnathostomes or stem vertebrates, close to but possibly outside the crown group of vertebrates.2, 15
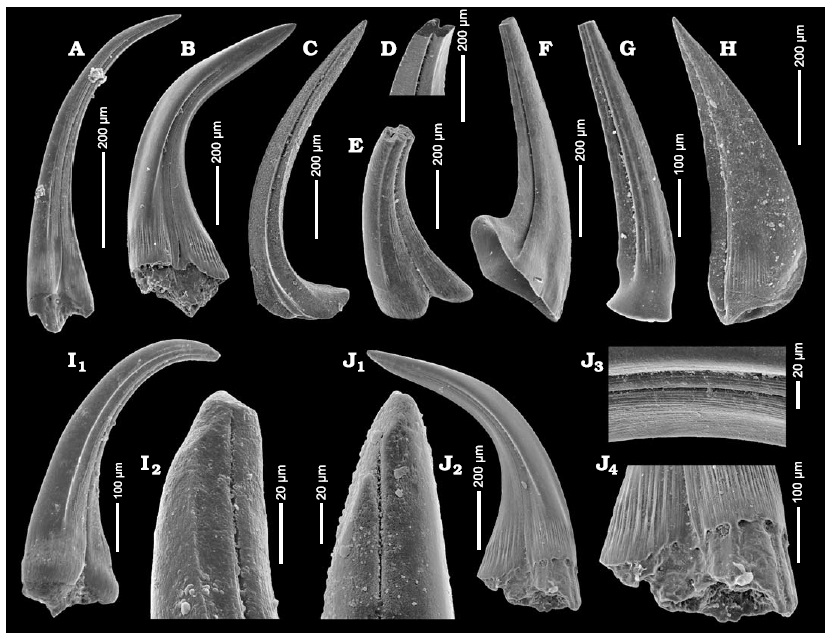
Element morphology and apparatus structure
Conodont elements occur in three main morphological categories: coniform (simple cone-shaped), ramiform (with one or more processes bearing denticles), and pectiniform (platform-like elements with broad surfaces).6, 13 In life, these elements were arranged in a bilaterally symmetric apparatus within the head region, typically comprising 15 to 19 elements organized into a series of paired and unpaired positions. The apparatus functioned as a feeding structure, with different element types performing different roles: platform elements may have served as grinding surfaces for food processing, while coniform and ramiform elements functioned in grasping and slicing.7, 8
Evidence for the feeding function of conodont elements comes from multiple lines of inquiry. Microwear analysis reveals scratches and pits on element surfaces consistent with tooth-on-tooth contact during food processing, and the patterns of wear vary among element positions in ways that match the predicted mechanical stresses of a functioning feeding apparatus.8 The elements grew by periodic accretion of new layers, and damaged elements show evidence of repair during life, further supporting an active functional role.6, 14
Growth and development
Conodont elements grew by the sequential addition of lamellae — thin layers of apatite deposited on the surface of the element. This incremental growth pattern is visible in cross-section and has been compared to the growth rings of trees, potentially recording periodic (possibly daily or tidal) cycles of mineral deposition.14 The growth pattern of conodont elements is fundamentally different from that of vertebrate teeth, which grow from within by progressive mineralization of a preformed organic matrix. In conodonts, growth occurred by surface accretion from surrounding soft tissue, a process more akin to the formation of dermal bone or scales.6, 14
Ontogenetic series of conodont elements show that element morphology changed significantly during growth, with juvenile elements often having simpler morphologies that became progressively more complex with each growth increment. This ontogenetic variation has been a source of taxonomic confusion, as growth stages of a single species have sometimes been described as separate taxa.6
Biostratigraphic utility
The biostratigraphic value of conodonts is exceptional. Conodont elements are extremely abundant in marine sedimentary rocks — a single kilogram of limestone may yield thousands of specimens — and they can be extracted efficiently by dissolving the rock in dilute acetic or formic acid, which destroys the carbonate matrix while leaving the phosphatic elements intact.13 Conodont species evolved rapidly, often with stratigraphic ranges of less than one million years, and many achieved wide or global distributions in marine environments. These properties make conodonts the primary biostratigraphic tool for Ordovician through Triassic marine sequences, surpassing graptolites, ammonites, and even trilobites in resolution for many intervals.13
Conodont-based biozones provide the framework for international correlation of Ordovician, Silurian, Devonian, Carboniferous, Permian, and Triassic stages, and several Global Boundary Stratotype Section and Points (GSSPs) are defined by first appearances of conodont species.13
The conodont alteration index
Beyond biostratigraphy, conodont elements serve as geothermometers through the conodont alteration index (CAI), developed by Epstein, Epstein, and Harris in 1977.12 When heated, the originally pale-colored apatite of conodont elements progressively darkens through a sequence of color changes — from pale yellow through brown, dark brown, black, gray, and ultimately white (at very high temperatures) — caused by the thermal maturation of organic matter trapped within the crystal lattice. The CAI scale ranges from 1 (unaltered, less than 50°C) to 5 (fixed carbon, 300–480°C), with higher values assigned to elements that have been further altered by incipient recrystallization.12 Because the color change is irreversible and dependent on both temperature and duration of heating, the CAI provides a measure of the maximum thermal exposure experienced by the host rock, making it valuable for assessing the thermal maturity of petroleum source rocks and the metamorphic history of sedimentary sequences.12
Paleoecology and trophic diversity
Although all known conodont body fossils indicate small, eel-like animals, the diversity of element morphologies across conodont lineages suggests a wide range of feeding ecologies. Conodonts with robust platform elements bearing denticulate surfaces are interpreted as macrophagous processors that could crush or shear relatively large food particles, while those with delicate, comb-like ramiform elements may have been suspension feeders or microphagous grazers that raked particles from the water column or substrate.7, 20 The Ordovician radiation of conodonts produced an explosive diversification of element morphologies, paralleling the broader Great Ordovician Biodiversification Event and suggesting that conodonts were partitioning trophic niches in increasingly complex marine ecosystems.17
Oxygen isotope ratios extracted from conodont apatite have also been used as a paleothermometer for Paleozoic ocean temperatures. Because the oxygen isotopic composition of apatite is temperature-dependent at the time of mineralization, and because conodonts grew their elements incrementally throughout life, conodont apatite records the ambient water temperature of the environment in which the animal lived. Joachimski and colleagues demonstrated that conodont oxygen isotope data can resolve temperature fluctuations on timescales of less than one million years, providing a high-resolution paleoclimate proxy for intervals where other temperature proxies are sparse or absent.18 This application has been particularly valuable for documenting temperature shifts associated with the late Devonian and end-Permian extinction events, where conodont-based temperature curves reveal warming pulses coincident with biodiversity declines.18, 19
The Late Triassic decline of conodonts was not abrupt. Conodont diversity had been waning since the Carnian Stage (approximately 237–227 Ma), with progressive reductions in both species richness and morphological disparity. Rigo and Joachimski linked this decline to the Carnian Pluvial Episode, a period of climatic upheaval involving increased rainfall, ocean stratification, and reduced carbonate production that may have disrupted the marine ecosystems on which conodonts depended.19 By the Rhaetian Stage (the final stage of the Triassic), only a handful of conodont species persisted, making the group highly vulnerable to the environmental perturbations that accompanied the end-Triassic extinction.16, 19
Extinction
Conodonts survived four of the “Big Five” mass extinctions — the end-Ordovician, late Devonian, end-Permian, and Triassic–Jurassic events — suffering significant losses each time but recovering to diversify anew. They did not survive the end-Triassic extinction, however: the last known conodonts disappear at or near the Triassic–Jurassic boundary, approximately 201 million years ago, and the group left no descendants.13, 16 The cause of their final extinction is debated but is generally attributed to the same environmental perturbations — volcanism associated with the Central Atlantic Magmatic Province, ocean acidification, and marine anoxia — that drove the broader end-Triassic extinction event.16