Overview
- Human language rests on a suite of biological adaptations including a permanently descended larynx, fine motor control of respiration, enlarged Broca's and Wernicke's areas, and a derived form of the FOXP2 gene that is essential for the precise orofacial movements speech requires.
- Archaeological evidence for symbolic behavior—pigment use, shell beads, engraved geometric patterns, figurative art—converges on a period between 100,000 and 50,000 years ago in Africa and suggests that fully modern language capacity was in place by the time of the dispersal out of Africa.
- Of roughly 7,000 languages spoken today, more than half are expected to fall silent by 2100; the field of historical linguistics uses the comparative method to reconstruct ancestral proto-languages, tracing language families back tens of thousands of years.
Language is the most distinctively human of all behaviors. No other species has developed a communication system of comparable complexity, productivity, or expressive range. Spoken human languages are generative in a mathematically precise sense: a finite inventory of sounds is combined according to grammatical rules to produce an effectively infinite number of sentences, the vast majority of which any speaker has never heard before.6 This capacity rests on a suite of biological prerequisites—anatomical, neurological, and genetic—that differentiate Homo sapiens from all other living primates. Yet language is also a cultural artifact: it is learned within communities, transmitted across generations, and subject to continuous change. The roughly 7,000 languages spoken in the world today are the products of tens of thousands of years of divergence from ancestral proto-languages, a process that historical linguists study using rigorous comparative methods.12, 14
The evolutionary origins of language remain one of the most debated questions in science. Direct evidence is scarce: spoken language leaves no fossil record, and the brain structures that support it are visible only in endocranial casts of limited interpretive power. Researchers therefore work with indirect proxies—the anatomy of the vocal tract, the genetics of language-related genes, the neuroscience of speech production and comprehension, and the archaeology of symbolic behavior—to reconstruct when and how language emerged in the hominin lineage.6
The biological prerequisites for speech
Speech, as opposed to language in the abstract sense, depends on precise neuromuscular control of the vocal tract: the larynx, pharynx, tongue, lips, velum, and the muscles of respiration. In humans, the larynx sits lower in the neck than in other adult primates, a configuration that enlarges the supralaryngeal vocal tract and permits the production of a broader range of formant frequencies, including the vowels that carry much of the phonemic contrast in human languages.3 This descended larynx was long considered uniquely human, but research by Fitch and Reby demonstrated in 2001 that other mammals, including red deer, also temporarily lower the larynx during vocalization, suggesting that the anatomical configuration in humans is an extreme and permanent elaboration of a more widespread mammalian capacity rather than a wholly novel structure.3 The descended larynx also increases the risk of choking, a hazard that has no parallel in other apes, which suggests that the benefits to communication must have been substantial enough to be favored by natural selection despite this cost.6

{kind=link}
Fine voluntary control of breathing is equally essential. Human speakers actively regulate subglottal air pressure during speech, modulating it phrase by phrase in ways that no other primate can match. The neural pathway responsible for this control involves direct cortical projections to the spinal motor neurons that drive the diaphragm and intercostal muscles. In non-human primates, this direct corticospinal pathway is absent or greatly reduced; vocalizations are largely controlled by subcortical structures and are stereotyped and involuntary in character.6
Two cortical regions are of central importance to language in humans. Broca's area, in the left inferior frontal gyrus, is involved in speech production, grammatical processing, and the sequencing of linguistic units. Wernicke's area, in the left posterior superior temporal gyrus, is critical for the comprehension of spoken language. These regions are connected by the arcuate fasciculus, a white-matter pathway that has undergone significant elaboration in humans compared to other great apes. Imaging studies have shown that comparable regions exist in chimpanzees and other primates, but they are substantially smaller relative to overall brain size and appear to serve primarily gestural and auditory discrimination functions rather than the syntactic processing characteristic of human speech.6
The FOXP2 gene and the genetics of speech
The identification of the FOXP2 gene provided the first specific molecular insight into the genetic basis of human language. Mutations in FOXP2 were identified in 2001 as the cause of a severe and specific language disorder in a large British family (the KE family) spanning three generations. Affected members of this family suffer from difficulties with orofacial movements required for speech, impaired grammatical processing, and deficits in both language production and comprehension, despite having otherwise normal intelligence.21 FOXP2 encodes a transcription factor of the forkhead box family, meaning it regulates the expression of many downstream genes; it is expressed in the brain, lungs, heart, and gut during development.1
In 2002, Wolfgang Enard and colleagues published a comparative analysis of FOXP2 across mammals. They found that the protein-coding sequence of FOXP2 is highly conserved across mammals generally, but that the human lineage had accumulated two amino acid substitutions absent in all other examined mammals. Population genetic analysis revealed signatures of a selective sweep in the region surrounding these substitutions in humans, indicating that the derived human form of FOXP2 was strongly favored by natural selection relatively recently in evolutionary history—within the past 200,000 years.2 Subsequent work identified that the FOXP2 gene in mice, when its two human-specific mutations were introduced by knock-in techniques, produced mice with altered ultrasonic vocalizations and modified basal ganglia circuitry, confirming the gene's causal role in the motor learning and auditory-motor integration that underpin vocal behavior.20
FOXP2 is not a "language gene" in the sense of encoding language itself; rather, it regulates a developmental program that shapes the neural circuits involved in the learning and production of complex, precisely timed motor sequences, of which speech is one example. Birds that learn songs (a capacity analogous to human speech learning, though independently evolved) also show elevated FOXP2 expression in song-learning regions of the brain during the sensitive period for song acquisition, underlining the gene's conserved role in vocal learning.20 Fisher and Scharff summarized the emerging picture in 2009: FOXP2 is a deeply conserved regulator of sensorimotor circuits, and the two human-specific amino acid changes refined its function in ways that contributed to the vocal learning ability underpinning speech.20
Did Neanderthals speak?
The question of whether Neanderthals possessed language-like communication is one of the most contested in paleoanthropology. Evidence bears on it from several directions: the anatomy of the Neanderthal vocal tract, their genome, and the archaeological record of their behavior. In 1983, excavations at Kebara Cave in Israel recovered a remarkably complete Neanderthal hyoid bone—the small U-shaped bone in the throat to which the muscles of the tongue and larynx attach—dating to approximately 60,000 years ago. Arensburg and colleagues reported in 1989 that the Kebara hyoid is morphologically indistinguishable from that of modern humans, in contrast to the hyoids of chimpanzees and other non-human primates, which differ substantially in shape and position.4 This finding has been interpreted by some researchers as evidence that the Neanderthal larynx was positioned in a manner compatible with speech, though others have cautioned that hyoid shape alone does not determine vocal tract configuration.5
Biomechanical analysis of Neanderthal ear ossicles has provided further anatomical evidence. Studies of the bony labyrinth of Neanderthal temporal bones show that their hearing range was biased toward the same frequencies most critical for speech intelligibility in modern humans, suggesting that auditory anatomy was adapted to process speech-like signals even if the full speech apparatus remains debated.5 Computational reconstructions of the Neanderthal supralaryngeal vocal tract by Barney and colleagues suggested that Neanderthals were capable of producing a broad range of vowel-like sounds, though possibly with somewhat different formant patterns than modern humans.5
The genomic evidence has been particularly illuminating. Sequencing of ancient DNA from Neanderthal fossils revealed that Neanderthals carried the same two derived amino acid substitutions in FOXP2 that are found in all modern humans and absent in chimpanzees.22 This finding demonstrates that the molecular substrate for the fine orofacial motor control that speech requires was present in Neanderthals. However, a 2013 analysis by Maricic and colleagues identified a regulatory region near FOXP2 that differs between Neanderthals and modern humans, suggesting that despite sharing the same protein sequence, FOXP2 expression may have been slightly different between the two species.22 The consensus that has emerged is cautious: Neanderthals almost certainly had some form of complex vocal communication, but whether it constituted full language with syntax comparable to that of modern humans remains unresolved.
Theories of language origin
Several major hypotheses address how language first evolved in the hominin lineage, and the field has cycled through these debates repeatedly since the subject became scientifically tractable in the late twentieth century. A persistent division exists between those who argue that language evolved primarily from gestural systems and those who favor a vocal origin.
The gestural hypothesis, in its modern form, draws on the discovery of mirror neurons in the macaque premotor cortex—neurons that fire both when an animal performs an action and when it observes another individual performing the same action. Rizzolatti and Arbib proposed in 1998 that the mirror neuron system provides a neural substrate for linking observation, imitation, and intentional communication, suggesting that gestural communication evolved first and was subsequently accompanied by vocalizations that became increasingly structured and mandatory over evolutionary time.8 Arbib elaborated this into the "mirror system hypothesis" for language evolution, arguing that Broca's area in humans is a direct evolutionary descendant of the macaque mirror neuron region, and that the capacity for imitation enabled by this system was a prerequisite for the cultural transmission of linguistic conventions.7 Comparative evidence supports a significant role for manual gesture in human language: gestures and speech are co-produced in normal communication, deaf communities spontaneously develop fully structured sign languages that recruit the same neural networks as spoken language, and children exposed to both gesture and speech show gesture systematically leading speech development in semantic complexity.6
The vocal grooming hypothesis, proposed by Robin Dunbar, suggests that language evolved as a more efficient replacement for the physical social grooming that maintains bonds in primate societies. As hominin group sizes increased beyond what physical grooming could sustain, vocal grooming—eventually elaborating into language—allowed individuals to manage a larger network of social relationships simultaneously.6 A related perspective, sometimes called the ritual or music hypothesis, draws attention to the role of rhythmically coordinated vocalizations—shared songs, chants, and group sound-making—as precursors to referential language. Steven Mithen developed this view in his 2005 synthesis The Singing Neanderthals, arguing for an ancestral "Hmmmmm" communication system (holistic, manipulative, multi-modal, musical, and mimetic) that predated the compositional and referential features of modern language.19
No single hypothesis commands consensus, in part because different aspects of language—phonology, syntax, semantics, pragmatics—may have evolved at different times and through different selective pressures. Most researchers now regard language as a mosaic adaptation, with different components co-opting and elaborating upon pre-existing neural systems in parallel rather than evolving as a single integrated package at one point in time.6
The archaeological evidence for language
Because spoken language leaves no physical trace, archaeologists use the appearance of symbolic and abstract thought as a proxy for language capacity. The reasoning is that the cognitive operations required to assign arbitrary meanings to sounds or symbols—and to manipulate those symbols according to combinatorial rules—are the same operations that produce deliberate ornamentation, representational art, and long-distance exchange networks. Sites in southern Africa have produced some of the earliest evidence for this cognitive profile.
Blombos Cave on the southern Cape coast of South Africa has yielded a sequence of symbolic artifacts spanning the Middle Stone Age. Engraved ochre pieces bearing geometric crosshatch patterns date to approximately 75,000 years ago, representing the earliest firmly dated abstract engravings known anywhere in the world.11 An ochre-processing workshop at the same site, dating to around 100,000 years ago, included abalone shells used as mixing containers, containing a compound of ochre, bone, charcoal, and fat—evidence of multi-component recipe preparation requiring planning and symbolic intent.25 Perforated Nassarius shell beads from Blombos and other sites in North Africa and the Near East, some dating to 130,000–100,000 years ago, are interpreted as body ornamentation—the use of visible symbols to communicate social identity, a behavior that presupposes shared conventions of meaning.26
The Sibudu Cave site in KwaZulu-Natal has provided evidence of complex plant-based bedding and hafting compounds by approximately 77,000 years ago, indicating forward planning and knowledge transmission across generations.9 By 50,000–40,000 years ago, the Upper Paleolithic transition in Europe is marked by the appearance of figurative art, personal ornaments, and musical instruments (bone flutes) in a broad expansion of symbolic culture often associated with the dispersal of anatomically modern humans out of Africa, though recent finds of cave art at sites in Sulawesi dated to over 45,500 years ago by uranium-series dating demonstrate that symbolic behavior was not confined to Europe or to a single time horizon.10
The convergence of evidence from multiple regions and multiple behavioral domains supports the interpretation that fully modern language capacity was present in Homo sapiens by at least 100,000 years ago, and possibly substantially earlier.11, 25, 26 Whether earlier hominins such as Homo heidelbergensis or Neanderthals possessed rudimentary language remains an open question, though the anatomical and genomic evidence outlined above indicates they were not without relevant biological prerequisites.
Language families and linguistic diversity
The world's approximately 7,000 living languages—a figure documented by Ethnologue, the most comprehensive linguistic database—are grouped by historical linguists into families on the basis of shared ancestry, and many remain unclassified or are considered language isolates with no demonstrated relationship to any other language.12 The distribution of linguistic diversity is highly uneven: the Americas and the Pacific (particularly Papua New Guinea) account for the majority of the world's language families despite having far smaller populations than Eurasia, a pattern consistent with long periods of small-group isolation in linguistically diverse communities following the initial peopling of those regions.28
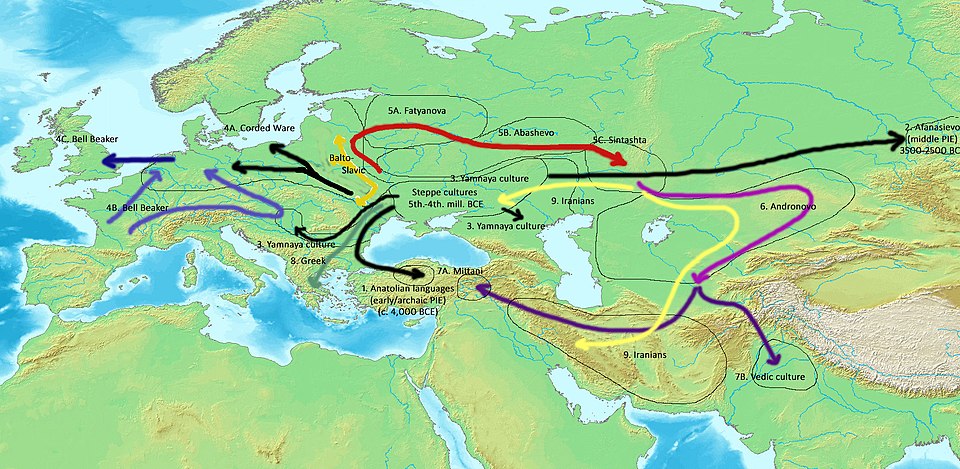
The Indo-European family is the most extensively studied and encompasses languages from Ireland to Bangladesh, including the Germanic, Romance, Slavic, Indic, and Iranian branches. Phylogenetic analysis of lexical cognates by Gray and Atkinson in 2003 estimated that the Indo-European family began diverging approximately 8,700–9,800 years ago, consistent with the Anatolian hypothesis that links Proto-Indo-European with the spread of early farming from Anatolia into Europe.15
{kind=link}
A subsequent Bayesian phylogeographic analysis by Bouckaert and colleagues in 2012 confirmed this timing and origin using a larger dataset, mapping the diffusion of daughter languages across the continent over millennia.16 The Sino-Tibetan family, which includes Mandarin Chinese, Tibetan, and Burmese, is the second largest by number of speakers and is thought to have begun diversifying roughly 4,000–7,000 years ago, though the deep internal structure of the family remains debated. The Niger-Congo family of sub-Saharan Africa encompasses over 1,500 individual languages, making it the largest family by number of constituent languages; its best-known branch, Bantu, expanded across central and southern Africa within the past 5,000 years, a demographic event traceable in both linguistic and genetic data.27 The Austronesian family, extending from Madagascar to Easter Island and encompassing over 1,200 languages, represents one of the most geographically expansive colonizations in human prehistory, with Proto-Austronesian reconstructed to approximately 5,500 years ago in Taiwan.12
Selected major language families by approximate speaker count and geographic range12, 28
| Family | Approx. languages | Approx. speakers | Primary geographic range |
|---|---|---|---|
| Indo-European | 449 | 3.3 billion | Europe, South & Central Asia, Americas |
| Sino-Tibetan | 459 | 1.4 billion | East & Southeast Asia |
| Niger-Congo | 1,540+ | 700 million | Sub-Saharan Africa |
| Afro-Asiatic | 375 | 500 million | North Africa, Horn of Africa, Middle East |
| Austronesian | 1,250+ | 350 million | Southeast Asia, Pacific, Madagascar |
| Dravidian | 86 | 220 million | South India, Sri Lanka |
| Turkic | 41 | 170 million | Central Asia, Anatolia |
The comparative method and proto-languages
Historical linguistics reconstructs the ancestral languages from which attested language families descend through the comparative method, a systematic procedure for inferring sound correspondences between related languages and working backward to reconstruct earlier phonological, morphological, and lexical forms. The method was formalized in the nineteenth century by Indo-European scholars who recognized that systematic phonological correspondences between Sanskrit, Greek, Latin, Gothic, and Lithuanian pointed to a common ancestor language, now called Proto-Indo-European (PIE).14 A famous example is the word for "father": Sanskrit pitā, Greek patēr, Latin pater, Old English fæder—all reflecting a reconstructed PIE form *ph₂tér.14 The method does not rely on assumed regularities; instead, it discovers them empirically through the pattern of correspondences in the data. Where correspondences are regular and recurrent, common ancestry is the most parsimonious explanation.
The depth to which the comparative method can reach is limited by the rate of language change and the rate at which cognates are replaced by unrelated words. Estimates suggest that core vocabulary (numerals, body parts, basic verbs) changes at a relatively slow and approximately constant rate, enabling a rough molecular clock analogy for language—though this approach, called glottochronology, remains controversial because the rate of lexical replacement is known to vary by vocabulary domain, language community, and intensity of contact.14, 15 Most linguists consider reliable reconstruction impossible beyond approximately 8,000–10,000 years without exceptional documentary evidence, which is why proto-language proposals reaching further back in time—such as Joseph Greenberg's "Nostratic" macrofamily—remain speculative and contested.14
Language contact complicates reconstruction by introducing borrowed words and structural features that may obscure genetic relationships. Thomason and Kaufman documented in detail how intensive contact can lead to the wholesale restructuring of a language's grammar, not merely its vocabulary—a process called "metatypy" in extreme cases—blurring the boundaries between inherited and borrowed material.23 Despite these complications, the comparative method remains the most reliable tool available for inferring linguistic prehistory, and its results converge in important ways with independent evidence from archaeology and population genetics about the movements and interactions of ancient human groups.15, 16
The independent invention of writing
Writing systems represent a qualitatively distinct development in the history of language: the externalization of linguistic information into durable physical marks that can be transmitted across space and time without a speaking human as intermediary. Writing was not a simple invention made once and diffused globally; the archaeological record indicates that full writing systems developed independently in at least three, and arguably four, separate traditions.

{kind=link}
The earliest fully developed writing system known to archaeology is Sumerian cuneiform, which emerged in Mesopotamia around 3100 BCE. It originated in a system of clay tokens used for accounting in temple economies; Denise Schmandt-Besserat's influential analysis traced a developmental sequence from three-dimensional tokens through envelope sealing to impressed tablets to incised signs, showing how writing grew out of administrative necessity rather than literary aspiration.17 Egyptian hieroglyphs appear contemporaneously, around 3200–3100 BCE, and may reflect some stimulus diffusion from Mesopotamia—that is, awareness that writing existed without direct borrowing of the Sumerian system—though the Egyptian script operates on entirely different principles (logographic and phonetic rebus) and must be considered a substantially independent creation. Chinese writing, documented in oracle bone inscriptions from the Shang dynasty around 1200 BCE, shows no demonstrable connection to either Sumerian or Egyptian traditions and is regarded as fully independent; its antecedents may extend back to Neolithic potters' marks from around 5000 BCE, though the continuity between these marks and later script remains debated.18 Mesoamerican writing, attested in Olmec and Zapotec traditions by approximately 900–600 BCE and later elaborated in Maya glyphic script, represents yet another independent tradition with no contact relationship to Old World systems.17
The repeated independent invention of writing across unconnected cultures demonstrates that the cognitive prerequisites for symbolic notation—already present in language itself—readily give rise to visible notation systems when social complexity and administrative demands create sufficient pressure for their development. Conversely, the fact that writing emerged so late relative to the probable antiquity of spoken language (tens of thousands of years after the likely origin of modern language) underscores that writing is not a biological adaptation but a cultural technology built on top of an already-existing linguistic faculty.6
Language change and endangerment
All living languages change continuously. Sound systems shift, grammatical structures are reorganized, vocabularies expand through borrowing and coinage and contract through obsolescence. The linguist William Labov documented in detail how sound changes propagate through speech communities, showing that new pronunciations typically begin with specific social groups and spread across a community through social networks, often below the level of conscious awareness.24 Over centuries, such changes accumulate to the point where related languages become mutually unintelligible; over millennia, they diverge to the point where the relationship is detectable only through the formal application of the comparative method.14
Of the approximately 7,000 languages currently spoken, a substantial proportion face extinction within the next century. Crystal estimated that roughly half of the world's languages may disappear by 2100 if current trends continue, with one language dying every two weeks on average.13 Language death is accelerated by political marginalization of minority communities, economic incentives favoring dominant national or global languages, and the disruption of intergenerational transmission through migration and assimilation. The loss of a language entails the permanent loss of a unique classificatory system for natural phenomena, a body of oral literature and traditional ecological knowledge, and an irreplaceable record of human cognitive diversity.13
Approximate distribution of the world's ~7,000 languages by region28
Documentation and revitalization efforts by linguists, working alongside speaker communities, have produced grammars, dictionaries, and text corpora for many endangered languages, preserving structural and lexical data even when the number of fluent speakers has fallen to the point where community use is no longer viable. The scientific and humanistic importance of this work is substantial: each language encodes a distinct model of the world, and the loss of linguistic diversity diminishes humanity's collective record of the range of cognitive solutions that language permits.13
The development of language—from its biological underpinnings in early Homo to the diversification of thousands of language families, the invention of writing, and the ongoing dynamics of language change and loss—is one of the longest and most consequential processes in the natural and cultural history of the human species. It is simultaneously a chapter in evolutionary biology, in cognitive neuroscience, in archaeology, and in cultural history, and it can be fully understood only by drawing on all of these disciplines in concert.6, 14
References
Neanderthal-derived FOXP2 in modern humans is associated with altered brain activity in dorsal striatum
Phosphatic duricrust and the early appearance of complex cognition: shell beads from Skhul and Oued Djebbana