Overview
- Ornithischia, the 'bird-hipped' dinosaurs, encompassed all herbivorous dinosaur lineages outside Sauropodomorpha, including armored thyreophorans, duck-billed hadrosaurs, horned ceratopsians, and dome-headed pachycephalosaurs, united by a distinctive predentary bone and a retroverted pubis.
- Despite their name, ornithischians are not ancestral to birds—that lineage belongs to saurischian theropods—but they independently evolved a pubic arrangement convergent with that of modern birds, and their feeding innovations, including dental batteries with hundreds of interlocking teeth, represent some of the most sophisticated chewing systems in vertebrate history.
- Ornithischians dominated Late Cretaceous herbivore communities worldwide, with hadrosaurs and ceratopsians alone comprising more than half of large-bodied herbivorous dinosaur diversity in many Campanian and Maastrichtian assemblages before their extinction 66 million years ago.
Ornithischia, one of the two great divisions of Dinosauria, encompasses a spectacularly diverse radiation of herbivorous dinosaurs that shaped terrestrial ecosystems from the Late Triassic through the end of the Cretaceous Period.1, 2 The name, meaning "bird-hipped," refers to the superficial resemblance of the ornithischian pelvis to that of modern birds, in which the pubis is rotated posteriorly to lie roughly parallel to the ischium—a condition that evolved independently from the similar pelvic arrangement found in birds, which are actually descended from saurischian theropods.1, 7 Ornithischians include some of the most recognizable dinosaurs ever discovered: the plated Stegosaurus, the tank-like ankylosaurs, the horned ceratopsians such as Triceratops, the crested hadrosaurs, and the dome-headed pachycephalosaurs.2, 18 United by a suite of shared anatomical features—most notably the predentary bone at the tip of the lower jaw and a retroverted pubis—ornithischians evolved an extraordinary range of body plans, cranial ornaments, defensive structures, and feeding mechanisms over approximately 165 million years of evolution before all non-avian lineages perished in the end-Cretaceous mass extinction 66 million years ago.1, 3
Defining features and anatomy
Ornithischians are diagnosed by a constellation of skeletal features that distinguish them from saurischian dinosaurs. The most distinctive of these is the predentary, a small, unpaired bone at the anterior tip of the lower jaw that forms the bony core of the keratinous beak used for cropping vegetation.2, 5 No other vertebrate group possesses a predentary, making it a unique synapomorphy of Ornithischia.3, 5 The evolutionary origin of this bone has been clarified by the discovery of Sacisaurus, a Late Triassic dinosauriform from Brazil whose lower jaw bears paired elements at its symphysis that may be homologous to the ornithischian predentary, suggesting that the structure evolved from pre-existing components of the mandibular symphysis rather than arising as a completely novel element.5
The opisthopubic pelvis, in which the pubic bone is rotated posteriorly, represents another hallmark of the group.1, 2 This pelvic configuration enlarged the abdominal cavity, an adaptation likely related to the accommodation of a longer gut necessary for fermenting plant material, since all ornithischians for which dietary evidence exists were obligate herbivores.2, 23 Additional ornithischian synapomorphies include the absence of gastralia (ventral rib-like bones), the presence of a palpebral bone projecting over the orbit, an increased number of sacral vertebrae providing additional support for the trunk, and leaf-shaped or otherwise specialized cheek teeth with denticulate margins well suited to processing vegetation.2, 3
Recent work on ornithischian respiratory anatomy, informed by a remarkably well-preserved Heterodontosaurus specimen, has revealed that ornithischians evolved a unique ventilatory system distinct from the pneumatized, air-sac-driven respiration of saurischian dinosaurs.21 While theropods and sauropods developed extensive postcranial skeletal pneumaticity connected to avian-style air sacs, ornithischians retained more conservative lung anatomy, relying instead on costal ventilation aided by gastralia-free abdominal flexibility, representing a fundamentally different evolutionary solution to the challenge of efficient gas exchange at large body sizes.21
Origins and early evolution
The earliest history of ornithischian dinosaurs remains one of the most poorly understood chapters in dinosaur evolution. Whereas saurischian dinosaurs are well represented in Late Triassic fossil assemblages from South America and elsewhere, unambiguous ornithischians are extremely rare before the Early Jurassic, creating what has been termed the "ornithischian gap" in the Triassic fossil record.4, 20 For decades, Pisanosaurus mertii from the Ischigualasto Formation of Argentina (approximately 229 million years old) was considered the earliest known ornithischian, but phylogenetic reassessments published in 2017 and 2018 have recovered Pisanosaurus within Silesauridae, a group of non-dinosaurian dinosauriforms, potentially removing ornithischians from the Triassic record entirely.6
If the silesaurid reclassification of Pisanosaurus is correct, then the oldest unambiguous ornithischians date to the Early Jurassic. Lesothosaurus diagnosticus, from the Upper Elliot Formation of South Africa (approximately 200 million years old), is a small, lightly built bipedal herbivore that represents one of the most basal well-known ornithischians.4 Similarly, Eocursor parvus, also from southern Africa, provides evidence that early ornithischians were small-bodied, cursorial animals with relatively simple dentitions.4 The rarity of Triassic ornithischians contrasts sharply with the relative abundance of early saurischians, suggesting either a genuine late origin for the clade, a significant preservational or ecological bias against their fossilization, or the possibility that silesaurids represent stem ornithischians that would push the origin of the lineage back into the Middle Triassic.5, 20
The relationship between silesaurids and ornithischians has become one of the most active areas of research in dinosaur phylogenetics. Under the Ornithoscelida hypothesis proposed by Baron, Norman, and Barrett in 2017, ornithischians are the sister group of theropods rather than of sauropodomorphs, which would fundamentally restructure dinosaur phylogeny.7 Some analyses within this framework recover silesaurids as a paraphyletic grade of taxa along the stem leading to Ornithischia, providing a stepwise morphological transition from basal dinosauriforms to crown ornithischians, including the gradual evolution of the predentary and the retroverted pubis.7, 20 However, the traditional topology placing Ornithischia as the sister clade to Saurischia retains broad support, and the debate remains unresolved.7
Major ornithischian groups
Ornithischians are divided into two principal clades: Thyreophora, the armored dinosaurs, and Neornithischia, which includes all remaining ornithischians.2, 3 The position of Heterodontosauridae—small, fanged herbivores known primarily from the Early Jurassic of southern Africa—has varied across analyses, with some recovering them as the most basal ornithischians outside of both Thyreophora and Neornithischia, and others placing them within Neornithischia.2, 20
Thyreophora comprises two major subclades: Stegosauria and Ankylosauria. Stegosaurs, characterized by dorsal plates and tail spikes, flourished primarily during the Late Jurassic, with Stegosaurus stenops from the Morrison Formation of western North America serving as the best-known representative.8 Ankylosaurs, encased in extensive dermal armor composed of keeled osteoderms and, in derived forms, bearing massive tail clubs, reached their greatest diversity in the Late Cretaceous.8, 9 A comprehensive phylogenetic analysis of Thyreophora published in 2023, incorporating 340 characters scored across 91 taxa, challenged the traditional division of ankylosaurs into Ankylosauridae and Nodosauridae, instead recovering four distinct ankylosaur clades: Ankylosauridae, Panoplosauridae, Polacanthidae, and Struthiosauridae.8
Neornithischia is further divided into Ornithopoda and Marginocephalia. Ornithopoda ranges from small, bipedal "hypsilophodontids" of the Jurassic to the large, often crested hadrosaurs of the Late Cretaceous, which were among the most abundant and diverse large herbivores of their time.2, 13 Marginocephalia unites two superficially dissimilar groups: Ceratopsia, the horned dinosaurs, and Pachycephalosauria, the dome-headed dinosaurs, linked by the shared possession of a bony shelf extending from the posterior margin of the skull.2, 18 Ceratopsians radiated dramatically in the Late Cretaceous, producing the massive-frilled and multi-horned chasmosaurines and centrosaurines that dominated western North American herbivore communities.18, 19
Major ornithischian clades and their temporal ranges2, 8, 18
| Clade | Major subgroups | Temporal range | Key features |
|---|---|---|---|
| Heterodontosauridae | — | Late Triassic?–Early Cretaceous | Caniniform tusks, heterodont dentition |
| Thyreophora | Stegosauria, Ankylosauria | Early Jurassic–Late Cretaceous | Dermal armor, plates, tail spikes/clubs |
| Ornithopoda | Iguanodontia, Hadrosauridae | Middle Jurassic–Late Cretaceous | Dental batteries, bipedal/facultatively quadrupedal |
| Ceratopsia | Psittacosauridae, Neoceratopsia | Late Jurassic–Late Cretaceous | Rostral bone, horns, neck frills |
| Pachycephalosauria | — | Late Cretaceous | Thickened skull dome, ornamented skull margin |
Armor, horns, and defense
Ornithischian dinosaurs evolved an unparalleled diversity of defensive and display structures. Thyreophorans developed extensive dermal armor composed of osteoderms—bony elements embedded in the skin—that ranged from flat scutes and keeled plates to elongated spikes and, in ankylosaurids, massive tail clubs formed by the fusion of distal caudal vertebrae with enlarged osteoderms.8, 9 The functional significance of thyreophoran armor has been debated extensively. In ankylosaurs, the dermal armor was clearly defensive in primary function, but histological analysis has revealed dense vascularization within osteoderms, suggesting that thermoregulatory roles comparable to those proposed for crocodilian osteoderms may have been secondarily important.9

.jpg){kind=link}
The dorsal plates of stegosaurs present a more complex interpretive challenge. The large, thin plates of Stegosaurus were extensively vascularized and had relatively low structural strength, making them poor candidates for defensive armor.8 The leading hypothesis holds that stegosaur plates functioned primarily in species recognition and intraspecific display, with thermoregulation as a possible secondary function enabled by the dense vascularity.8 The tail spikes, or thagomizers, were structurally robust and clearly functioned as defensive weapons, as evidenced by pathological theropod bones bearing puncture wounds consistent with stegosaur tail-spike impacts.8
Ceratopsian horns and frills represent a separate evolutionary solution to display and defense. The functional evolution of the ceratopsian head involved the progressive elaboration of the nasal horn, brow horns, and parietal-squamosal frill through ontogeny and across the clade.19 Pathological evidence from Triceratops squamosal bones demonstrates that the brow horns were used in intraspecific combat: lesion frequencies on the squamosal are approximately ten times higher in Triceratops than in the centrosaurine Centrosaurus, consistent with predictions from biomechanical models of horn-locking combat between individuals possessing long brow horns.10 In centrosaurines, which possessed a single enlarged nasal horn, combat may have involved flank-butting rather than horn-locking, and the elaborate frill ornamentation likely served primarily as a species-recognition signal and display structure.10, 19
Feeding innovations
Ornithischians evolved some of the most sophisticated herbivorous feeding mechanisms in vertebrate history, progressing from the simple leaf-shaped dentitions of basal forms to the highly derived dental batteries of hadrosaurs and ceratopsians.11, 13 Dental evolution across the clade followed a consistent directional trend: basal ornithischians possessed relatively simple, isodont dentitions with leaf-shaped teeth bearing coarse marginal denticles suited for basic mechanical reduction of plant material, while derived groups evolved increasingly complex occlusal surfaces, tightly packed tooth rows, and precise dental occlusion that enabled efficient oral processing of tough, fibrous vegetation.13, 23
The hadrosaurid dental battery represents the pinnacle of dental complexity among dinosaurs. Hadrosaur jaws contained hundreds of teeth arranged in tightly interlocking columns and rows, with new teeth continuously growing beneath functional ones and being pushed into occlusion as older teeth were worn away.12 Ontogenetic studies have revealed that a critical evolutionary innovation underlying the dental battery was the halting of normal tooth root resorption, which allowed tooth roots to become functionalized as part of the occlusal surface, greatly expanding the grinding area available for food processing.12 Dental microwear analysis indicates that hadrosaurs employed complex, three-dimensional jaw movements including a transverse (side-to-side) component that is otherwise rare among non-mammalian vertebrates, enabling them to process tough vegetation with an efficiency approaching that of mammalian mastication.22
Ceratopsians independently evolved their own form of dental battery, with vertically stacked, self-sharpening teeth that formed a continuous shearing surface. Unlike the grinding mechanism of hadrosaurs, ceratopsian jaws operated primarily through a palinal (fore-aft sliding) motion combined with an orthal (vertical) bite, producing a slicing action suited to processing tough, woody plant material.11, 19 The evolution of increasingly caudally directed jaw adductor muscles across ornithischian phylogeny represents a broader trend: mechanical advantage of the jaw apparatus progressively increased through the enlargement of the coronoid process and the lowering of the jaw joint, enhancing bite force and enabling more powerful food processing in derived taxa.11
Relative jaw mechanical advantage across ornithischian groups11
Social behavior and reproduction
Ornithischian dinosaurs provide some of the best evidence for complex social behavior among non-avian dinosaurs. Mass bonebeds containing hundreds to thousands of individuals of a single species have been documented for hadrosaurs, ceratopsians, and ornithopods, strongly suggesting gregarious behavior and herding.14 The most spectacular of these accumulations is the Maiasaura bonebed in the Two Medicine Formation of Montana, which preserves an estimated 10,000 individuals and provides evidence for catastrophic mass mortality of a large herd.14
Ornithischians also furnish the earliest compelling evidence for parental care in dinosaurs. The original description of Maiasaura peeblesorum (the "good mother lizard") documented colonial nesting sites in which nests were spaced approximately 7 meters apart—roughly one adult body length—and contained juveniles with incompletely ossified bones and heavily worn teeth, indicating that hatchlings remained in the nest for an extended period and were provisioned by adults.14 Subsequent discoveries have extended evidence for parental care to other ornithischian lineages: a Psittacosaurus specimen from the Early Cretaceous Yixian Formation of China preserves an adult in close association with 34 juveniles, interpreted as evidence for post-hatching group care or crèching behavior.15
The elaborate cranial ornamentation of many ornithischians—the crests of lambeosaurine hadrosaurs, the horns and frills of ceratopsians, the domes of pachycephalosaurs—is widely interpreted as having functioned in intraspecific visual signaling, species recognition, and sexual display.19 In lambeosaurine hadrosaurs, the hollow cranial crests were continuous with the nasal passages and may have served as resonating chambers for vocalization, adding an acoustic dimension to social communication.18 The high species diversity of ceratopsians and hadrosaurs in Late Cretaceous western North America, where multiple closely related species sometimes co-occurred in the same formation, may have been driven in part by the role of species-specific cranial ornamentation as a pre-mating isolation mechanism.16, 19

_2_(15419555991).jpg){kind=link}
Ecological dominance in the Late Cretaceous
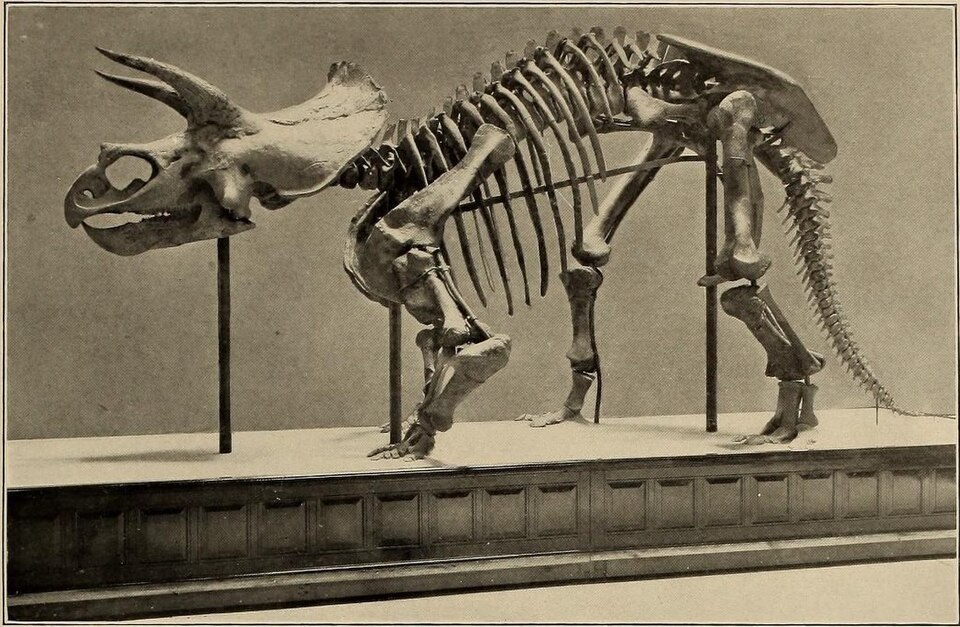
{kind=link}
Ornithischian dinosaurs reached their peak diversity and ecological dominance during the Late Cretaceous, particularly in the Campanian and Maastrichtian stages (approximately 84 to 66 million years ago).16, 17 In well-sampled terrestrial ecosystems such as the Dinosaur Park Formation of Alberta, Canada, ornithischians comprised the overwhelming majority of large-bodied herbivore diversity: hadrosaurs and ceratopsians together accounted for most of the megaherbivore biomass, with ankylosaurs present in lower abundances.16 Ecomorphological analysis of 14 megaherbivore genera from the Dinosaur Park Formation has demonstrated that contemporaneous hadrosaur and ceratopsian taxa occupied distinct regions of ecomorphospace, maintaining ecological separation through differences in skull shape, body size, and inferred feeding height and diet selectivity, a pattern that persisted through approximately 1.5 million years of species turnover in the formation.16
The co-radiation of ornithischians and angiosperms (flowering plants) during the Cretaceous has been hypothesized as a key driver of ornithischian diversification. The Cretaceous Terrestrial Revolution, as characterized by Lloyd and colleagues, involved a fundamental restructuring of terrestrial ecosystems driven by the radiation of angiosperms and the co-diversification of herbivorous insects, pollinating insects, and herbivorous dinosaurs.17 The timing of major ornithischian radiations—hadrosaurs diversifying in the Santonian to Campanian, ceratopsians in the Campanian to Maastrichtian—broadly corresponds to the period of angiosperm ecological dominance, though the precise causal links between plant and dinosaur diversification remain debated.17
Ornithischian biogeographic patterns in the Late Cretaceous reflect the progressive fragmentation of Pangaea and the formation of distinct landmasses with increasingly endemic faunas. Hadrosaurids achieved a nearly cosmopolitan distribution, reaching North America, South America, Europe, Asia, Africa, and possibly Antarctica, while ceratopsians remained largely restricted to North America and Asia, and ankylosaurs maintained a primarily Laurasian distribution with limited Gondwanan representation.24 This biogeographic partitioning produced regionally distinct ornithischian assemblages in the latest Cretaceous, with western North American faunas dominated by ceratopsids and hadrosaurids, Asian faunas featuring distinctive neoceratopsians and hadrosaurids, and European island faunas characterized by relictual and often dwarfed ornithopods and nodosaurids.24
Extinction and legacy
All ornithischian dinosaurs perished in the end-Cretaceous mass extinction approximately 66 million years ago, alongside all other non-avian dinosaurs.1 Unlike the saurischian theropods, which left avian descendants that survived the extinction boundary and radiated into the approximately 10,000 species of living birds, ornithischians produced no surviving lineages.1, 7 The complete extinction of Ornithischia is particularly striking given that the clade was at or near its peak diversity in the final stages of the Cretaceous: hadrosaurs and ceratopsians were the dominant herbivores in many terminal Maastrichtian ecosystems, and there is little evidence that ornithischian diversity was declining prior to the Chicxulub impact event.16, 17
The ecological roles vacated by ornithischians were eventually filled by mammals during the Cenozoic, but the transition was neither immediate nor straightforward. The large-bodied herbivore niches that ornithischians had occupied for over 100 million years went unfilled for millions of years after the extinction, and the mammalian herbivore lineages that eventually arose—perissodactyls, artiodactyls, proboscideans—arrived at broadly similar adaptive solutions through independent evolutionary pathways.1 The dental batteries of hadrosaurs find functional analogs in the hypsodont (high-crowned) teeth of horses and other grazing mammals, and the defensive armor of ankylosaurs is paralleled by the dermal ossifications of glyptodonts, representing striking examples of convergent evolution driven by similar ecological pressures operating on fundamentally different body plans.9
Ornithischian dinosaurs remain central to paleontological research not only for the extraordinary diversity of their morphological adaptations but also for the insights they provide into herbivore community ecology, the evolution of complex social behavior in deep time, and the dynamics of large-scale evolutionary radiations. Ongoing discoveries, particularly from previously undersampled regions of Gondwana and from the Triassic and Early Jurassic where the ornithischian record remains sparse, continue to reshape understanding of when, where, and how this remarkable group of dinosaurs originated and diversified.20, 24
References
A primitive ornithischian dinosaur from the Late Triassic of South Africa, and the early evolution and diversification of Ornithischia
A Late Triassic dinosauriform from south Brazil and the origin of the ornithischian predentary bone
Phylogenetic reassessment of Pisanosaurus mertii Casamiquela, 1967, a basal dinosauriform from the Late Triassic of Argentina
The phylogenetic relationships and evolutionary history of the armoured dinosaurs (Ornithischia: Thyreophora)
How Triceratops got its face: an update on the functional evolution of the ceratopsian head
A new Heterodontosaurus specimen elucidates the unique ventilatory macroevolution of ornithischian dinosaurs